RELATIVE DIFFERENCES BETWEEN RAPTORIAL APPENDAGES
A short study was conducted while on Heron Island Research Station involving the collection, observation, experimentation and subsequent return of one specimen of Gonodactylaceus falcatus.
"Left hook, right hook: Comparing the strike of left and right raptorial dactyls in G. falcatus"
Yolana Kailichova, University of Queensland

Summary
The second maxilliped of the stomatopod is utilised by the animal as a powerful tool a multitude of daily functions including for conspecific fighting, predation, and the construction of its burrow from coral rubble or rock. Biomechanic studies of the strike of the stomatopod demonstrates one of the fastest reflexes of any animal (Patek et al., 2005). clocking speeds of 20ms-1and this, notably is against the resistance of water (Patek & Caldwell,2004; Patek et al., 2007). The impact of such blows has been measured to reach peak forces of 1,500 N or 55kgs-1. This is extraordinary, for an organism which measures only centimetres in length. Notably, these figures take into account solely the force of one appendage and do not account for the cumulative effects of the cavitation bubble that ensues frequently after an initial impact (Patek et al., 2007).
This cavitation force can measure almost double the force of the initial strike. Thus, these cumulative forces are so powerful that on average only 2-3 strikes are required to break open the hard shells of their prey (Caldwell & Dingle,1975). Yet, it has been observed that when presented with larger prey, mantis shrimp's are capable of inflicting multiple blows over the course of several hours (Patek, 2004). All of this has consequences for the degradation of the stomatopod's own striking appendage and is solved by its molting behaviour (Caldwell, 1969).
Introduction
Given a well-characterised knowledge of the biomechanics and extent of the force of the strike, the question arises as to whether stomatopods typically maintain an equal force of impact over the course of consequent strikes.
This study aims to quantitate relative differences in impact between repeated strikes in Gondactylaceus falcatus comparing left, right and combined strikes.
Materials and Methods
Photographs 1, 2, 3, 5 by Yolana Kailichova.
Photographs 6, 7, 8 by Gustavo Alves.
One specimen of Mantis shrimp was collected from the Shark Bay at the University of Queensland's Heron Island Research Station. The specimen was translocated into large containers and placed under flowing seawater. The specimen was later transferred to a glass aquarium and allowed to settle (>1 hour). Ten pieces of plasticine were fashioned into square blocks maximising edge sharpness. These plasticine blocks were then placed on the end of a pencil. The block and pencil were presented in front of the animal. The moment of impact by the mantis shrimp was characterised by a loud single ‘tack’ sound.
Once the block was struck, the creation of two imprints could be observed. These are known to be caused by the two points of contact with the meral edges of the mantis shrimp’s two raptorial claws (Patek et al., 2005). Blocks, once struck, were numbered 1 -10 then set aside for later analysis. The results of the imprint were assigned by writing a number on each plasticine block (1-10). Steps 2 and 3 were performed for either i) 10minutes ii) 10 times or iii) until the animal refused to strike (determined as after 3 minutes of no contact).
The results of the double imprint on each block were then measured using a micropipette containing tap water. For each imprint (left, right and total), the volume of water was measured three times. This wasto account for projected measuring error, particularly involving the meniscus of water. These results were then compared graphically to those of the other blocks.
Results
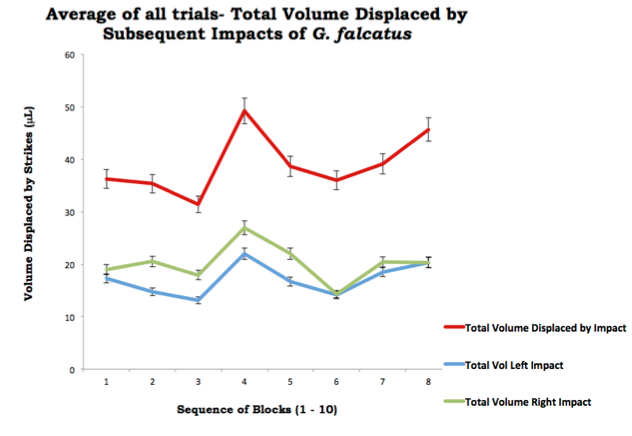
The results of the impact of each trial are outlined in the graph below. Error bars represent the replicate measurements for each trial. The final trial resulted in one initial strike followed by a period of 3 minutes with nocontact. Thus, only results from the first and second of the three trials were used in the final data set. For the initia ltwo trials, the peak of total volume from the two imprints measured 49.7μL, while the lowest volume generated was 31.35μL.
The general trend in the striking pattern of G. falcatus from trials 1 and 2 and their average, appeared to show a large imprint (as measured by the volume displaced) followed by two strikes of 'recovery' whereby lower results were recorded. The subsequent strike then peaked in volume (at 49.24μL) and was then followed by two lowervalues (38.63μL, 36μL respectively).
On the whole, the shape of the trend for the total, total left and total right volume, could be described as a sine curve.
While the data set is seemingly small, the values were consistent in this pattern. Interestingly, values for the right raptorial dactyl were consistently higher than those for the left (a peak of 26.97 and 22.0 for the right and left respectively). While this value is small, it is statistically significant.
Another interesting finding was for time lapsed between strikes. Time elapsed approximately doubled between impacts and greatly slowed as subsequent blocks were presented to the mantis shrimp. Block 1 took 0:12 seconds, while Block 1 in Trial 2 took 0:28 seconds, roughly doubling.
Discussion
The general trend of the graph, with a peak followed by two 'lower' impacts which build to a peak followed by one or two 'lower' strikes may be interpreted as fatigue, conditioning, or both. Stomatopods are well known for their high 'condition-ability' (Caldwell and Dingle; 1976) and researchers often rely on this attribute to examine their neurology and complex physiology (Marshall et al.,1996; Marshall et al., 1999). The general flattening out of the graph with the final two to three strikes may likewise reflect either motivation. Further, the increase in time elapsed between strikes, does little to illuminate the motivations behind the animal's strike, however the time lapsed between strikes may have higher explanatory value. Notably, after the fourth impact in the first trial, the specimen of G. falcatus swam away from the irritant. This behaviour was consistent between subsequent strikes.
There were several design limitations in the study, the result of limited time and resources and lack of experience. Some of these limitations included use of only one specimen, measurement error, subject conditioning, random angling of the block by experimenter and finally, use of coloured blocks with an animal possessing one of the most superior vision systems. This latter limitation was attempted to be resolved by conducting the experiment at night, however this may not have been sufficient. For future studies on impact, it would be suggested to utilise the experimental design of Patek & Caldwell (2005), whereby a static stimulus and highspeed videography is used.
Somewhat more conclusive is the consistent difference between left and right strike imprints, with the right hand creating a larger displacement volume than the left. Several animals have been known to display 'handedness'. Examples include heritable dexterity in wild chimpanzees' tool use (Forrester et al., 2012), the lateral coiling preferences of snakes (Roth, 2003) and the optical looking preferences of fish (Bisazza et al.,2000). However, should the results here prove to be replicable across the species, the congruency between the left and right results are perhaps less a case of 'handedness' stricto sensu than 'hand dominance'. Dominance, as measured by a greater realised force of an one side of an appendage over another, has been observed in Decapods such as fiddler crabs (Uca perplexa) and in this case, is largely driven by their morphological asymmetry (Claussen et al., 2008).
Interestingly, while the right appendage appeared to be slightly dominant to the left in any given strike, the cumulative effort produced was the same as the total average. This may say more about the recovery trend than about relative force applied, as measured between subsequent blocks. While the results are far from conclusive and may only speak for the one individual rather than the species G. falcatus, they do raise an interesting possibility of raptorial and or neurological asymmetry in mantis shrimps.
|