Chemoreception in the nudibranch, Platydoris scabra
Introduction
Mate searching can be particularly difficult in coral reefs due to the complex three dimensional environment and the high diversity of closely related species occurring in a close proximity (Little, Williams & Trowbridge 2009). Water is a viscous medium, as a result, locomotion in the marine environment is very energy expensive. Therefore in order to reduce energy expenditure, it is believed that strong selection is imposed on sensory systems to ensure that individuals can accurately detect potential mates. Chemical cues are frequently involved in the detection of preferred foods and mate searching in a variety of organisms including molluscs (Croll 1983; Susswein & Nagle 2004). To increase the likelihood of finding a mate, the gastropod Aplysia californica releases waterborne pheromones into the water column (Auderisk 1977). Gastropods are able to use their chemosensory organs to detect chemical cues (either released into the water column or incorporated into excretory fluids) and use these cues as an effective mate searching strategy. Platydoris scabra are frequently observed in pairs or aggregations of the same species. This behaviour has been identified as a prelude to mating in various opisthobranchs, with one individual often following a potential partner around for several hours (Karlsson & Haase 2002). I hypothesise that the release of chemical substances may facilitate the mate-searching behaviour in Platydoris scabra.
Chemoreception is the predominant sense employed by nudibranchs (Mollusca: Gastropoda) to perceive their environment (Harris 1971). The two main chemosensory structures in nudibranchs are the oral tentacles and rhinophores. Rhinophores are the primary chemosensory organs in nudibranchs and are primarily used to detect environmental cues (Murphy & Hadfield 1997). This study will begin by determining whether the cryptobranch dorid Platydoris scabra are more attracted to conspecifics (individuals of the same species) or to heterospecifics (individuals of a different species) based on the water soluble chemicals released from these individuals. Second, this study will investigate whether chemoreception is the primary method used by P. scabra to locate conspecifics and heterospecifics. This will be determined by placing a conspecific or a heterospecific into a plastic bag (to ensure no chemical cues are released into the water) and comparing the response of the P. scabra to these bagged individuals to the non-bagged individuals. This study will therefore determine whether chemoreception is the primary method used by P. scabra to differentiate between conspecifics and heterospecifics.
Method
Laboratory experiments were conducted at the Heron Island Research Station (HIRS), Australia, over a period of three days during September, 2012. The nudibranch P. scabra and the elephant slug Scutus antipodes (Image 1) were collected from beneath coral rubble on the reef crest at Heron Island. A total of 5 Platydoris scabra individuals were found (Image 2). One nudibranch was collected singly, meanwhile the others were found in pairs. The nudibranchs were enabled to acclimatize to the recirculating aquarium at HIRS for one day, and were utilized for the experiments on the second day after collection. Individuals were not fed throughout the experiment.

Image 1. Photograph of a Scutus antipodes specimen used throughout this experiment.

Image 2. The five Platydoris scabra speciments collected from Heron Island and used in this experiment.
The aim of this experiment was to quantify and compare the proximity of the subject to the conspecific (treatment 1) or heterospecific (treatment 2). The experimental set up consisted on 5 shallow plastic trays (28 cm wide and 40 cm long). Prior to each test, the trays were washed and sea water was replaced to remove any chemical cues present from the previous experiment. For both experiments, the test animals was placed in the centre of the tray (Figure 1). In experiment one, to eliminate the possibility of having two chemical stimuli, mucus cues were eliminated by placing either a conspecific or a heterospecific randomly in a corner of a tray (Figure 1). In experiment two, a conspecific (treatment 1) or a heterospecific (treatment 2) were placed into a plastic bag to remove any chemical cue, before being randomly placed in the corner of the tray (Figure 1). Each test lasted for approximately one hour, or until both individuals ceased movement. Each treatment was replicated with three individuals and each individual repeated the experiment thrice (n = 3). Each experiment was repeated twice on the first day (with a 5 hour break) and once on the following day to remove any learning effects. Photographs were taken at 5 minute intervals for the duration of each test. To determine the proximity of the subject to the conspecific or heterospecific a 6x5 grid was placed on top of the plastic tray. Individual subjects were scored based on their proximity to the heterospecific or conspecific (Figure 2). A value of 5 was awarded for the smallest distance between the subject and the heterospecific/conspecific, and a value of 0 was awarded for the maximun distance between these individuals. Tests where both of the animals did not move were not included. The data was analysed by a students paired, two-tailed t-test using the statistical software R.
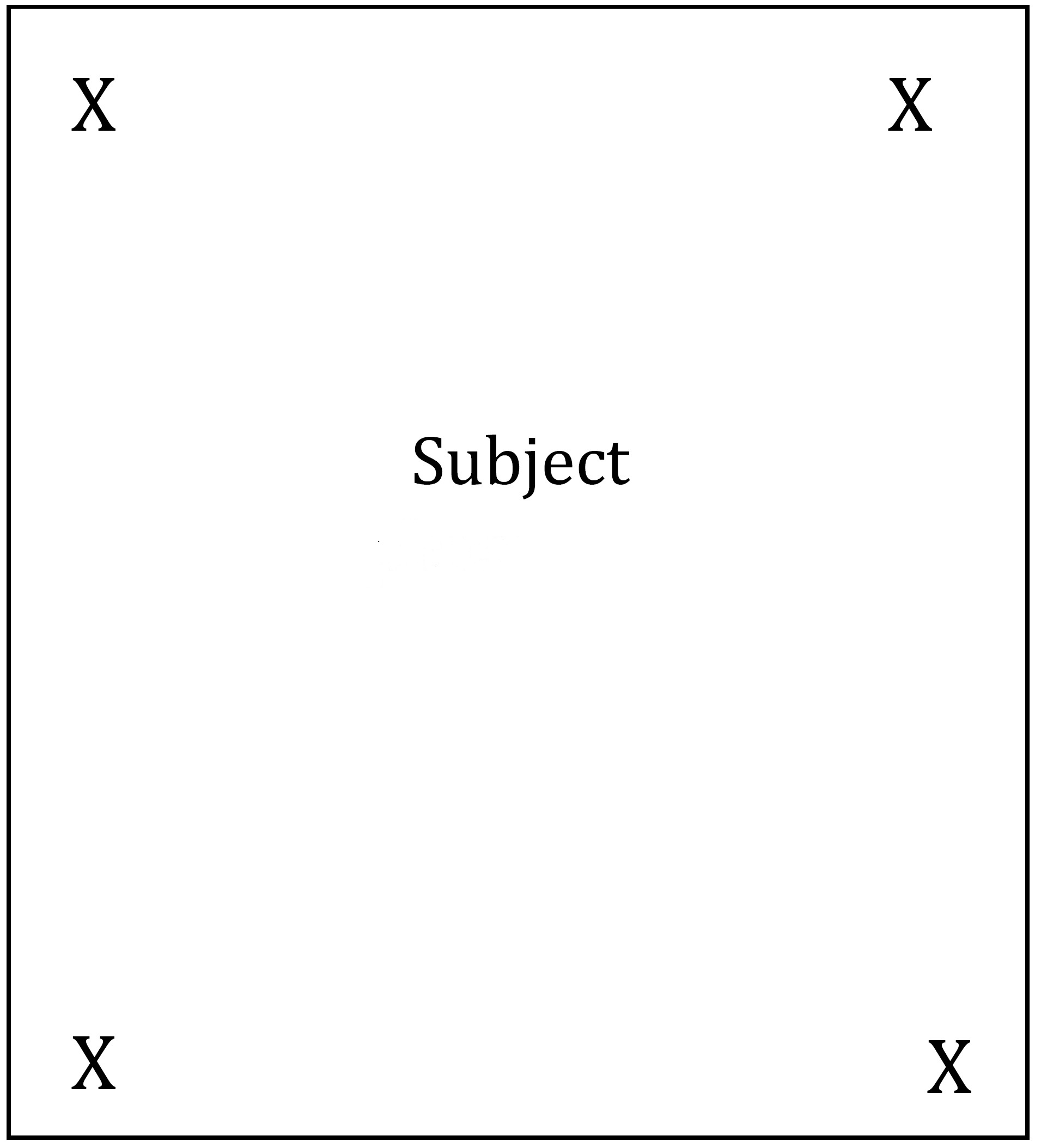
Figure 1. Experimental set up whereby the subject was placed in the centre of the tray and the conspecific/heterospecific was placed in a randomly designated corner (represented by the X's).
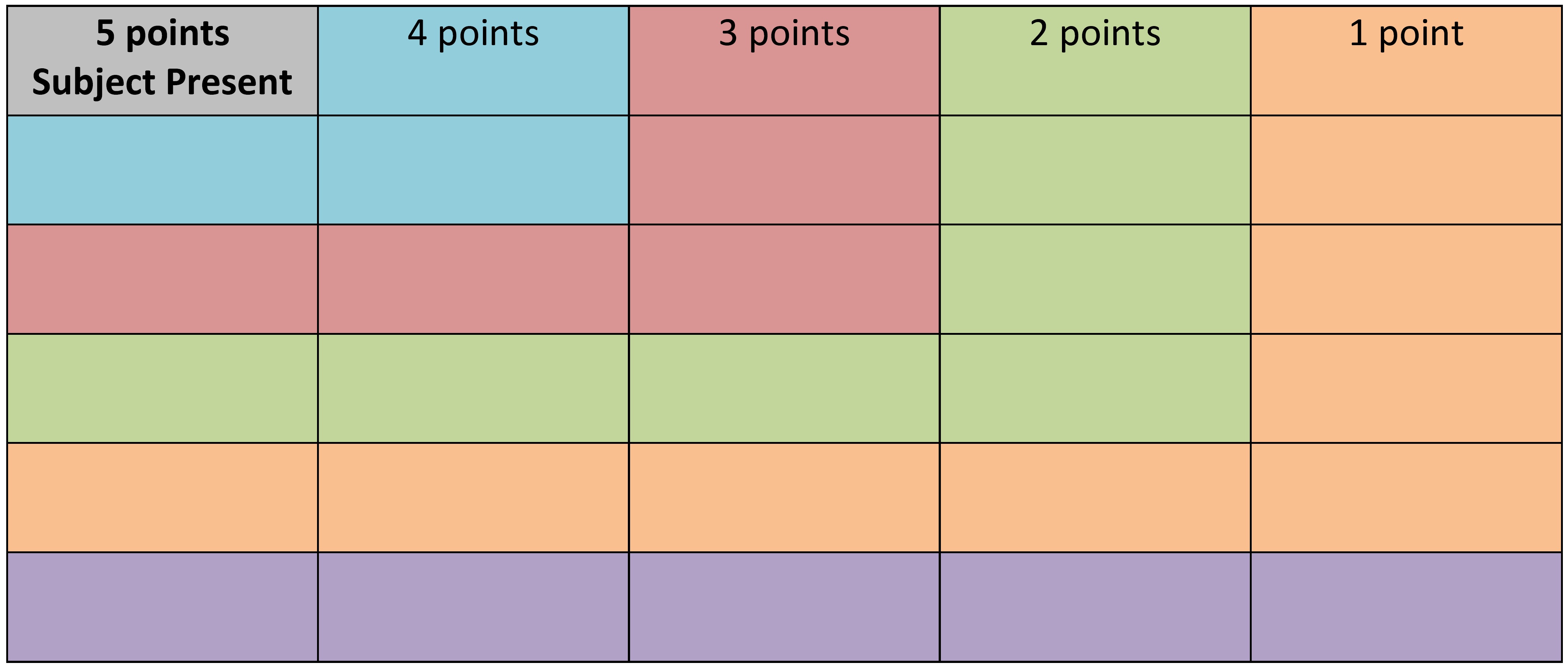
Figure 2. An example of the scoring system used to quantify the distance of the subject to the conspecific or heterospecific. If both the subject and the other individual were located in the same grid (represented by the grey square), they would score 5 points. However if the subject was located in the grey square and the other individual was located in the blue shaded area, they would score 4 points etc.
Results
In experiment one, all of the P. scabra subjects were significantly closer to conspecifics compared to heterospecifics (S. antipodes) after one hour (Figure 3, A, P < 0.05). In experiment two, no significant difference between P. scabra’s proximity to the bagged conspecifics and the bagged heterospecifics was observed (Figure 3, B, P > 0.05). There was very high variability in the distances between the subjects and the bagged individuals, with some individuals very close, and others very far away.
There was no significant difference between the proximity of the subject to the heterospecific without a bag and the heterospecfic within a bag (Figure 3, D, P > 0.05). When comparing the distance from the subject to the conspecific with, and without a platic bag, there was also no significant difference observed (Figure 3, D, P > 0.05).
One interesting observation noted during this study was that P. scabra initially moved towards both conspecifics and heterospecifics. Platyodoris scabra often touched or crawled over the top of these individuals (Figure 4, A). If P. scabra encountered a conspecific, they remained in close proximity to this individual. However, it was observed that if P. scabra encountered an heterospecific they quickly moved away from this individual (Figure 4, B).
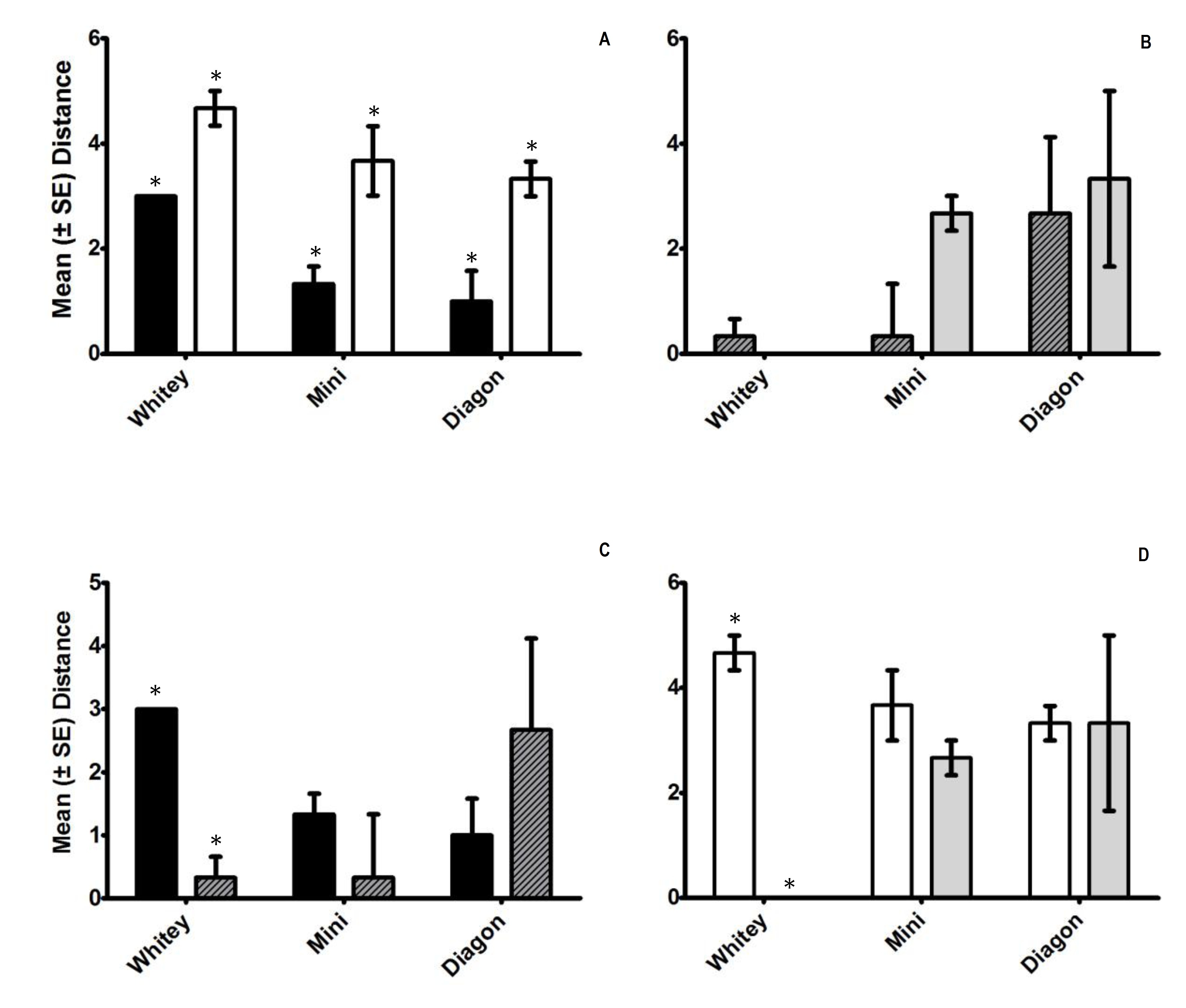
Figure 3. Mean ± SE proximity to Platydoris scabra (n=3). A value of 5 denotes the closest proximity to the subject, and a value of 1 denotes the greatest distance from the subject. Open bars denote conspecific treatments, black bars denote heterospecific treatments, striped bards denote that the heterospecifics were placed in a bag and grey bars denote that the conspecifics were placed in a bag. Asterisks indicate P < 0.05 (students paired, two-tailed t-test).

Figure 4. Observations for two trials conducted in experiment 1. (A) represents the subject (Platydoris scabra) with a conspecific. (B) represents the subject (Platydoris scabra) with a heterospecific (Scutus antipodes). Each image was taken at ten minute intervals (starting at time = 10 mins in the image on the left and concluding at time = 50 mins in the image on the right).
Discussion
Platydoris scabra occurred at a significantly closer proximity to conspecifics compared to heterospecifics (Figure 3, A). Therefore, it is likely that P. scabra were actively moving towards and remaining in close proximity with other individuals of the same species. This is likley due to the fact that they are potential mates. When the chemical cues were excluded by placing individuals in to a plastic bag, there was no significant difference in the distance between the subject P. scabra and the conspecifics and P. scabra and the subject and the heterospecifics. This suggests because there were no chemical cues the subject moved in a random direction, therefore having an equal possibility of either moving close to, or far away from the individual in the bag. From this study we can infer that P. scabra are more to occurr in aggregations of the same species rather than in aggregations with the sea slug S. antipodes. It is likely that P. scabra ae attracted to conspecifics (individuals of the same species) compared to heterospecifics (individuals of a different species) based on the water soluble chemicals released from these individuals.
The observations made during this study suggests that touch may also be an important mechanism for P. scabra to detect suitable mates. Platydoris scabra were initially attracted to both conspecifics and heterospecifics, however, once they touched the other individual with their aterior end, P. scabra were observed to move away from the heterospecifics and remain in close proximity of the conspecifics. The texture of P. scabra is very leathery compared to the texture of S. antipodes which is very soft (Image 1 and 2). Therefore, these observations suggest that the oral tentacles may be another important mechanism used to detect a suitable mate based on their texture. However, future work is necessary to investigate this explicitly.
Although we tried to eliminate the possibility of two chemical stimuli (by excluding mucus as a potential cue by placing individuals into the tray) once individuals were placed in this spot, there was no measure to ensure they did not move away from their initial locations. During multiple tests, conspecific individuals were observed to follow each other around the trays. Chemical compounds can be incorporated into excretory fluids (such as mucus) and many gastropods are able to use their chemosensory organs to follow mucus trails. This trail-following behaviour is widespread throughout gastropods and is an adaptive mate-searching strategy in the intertidal gastropod mollusc, Ilyanassa obsolete (Ng et al. 2011) and the nudibranchs Dendrodoris nigromaculata and D. nigra (Nakashima 1995). These nudibranchs can differentiate between conspecific and heterospecific mucous trails, only following the former, and detect directional information from mucous trails (Nakashima 1995). Dendrodoris nigromaculata and D. Nigra are similar to P. scabra as they are also predominantly feed on sponges and are observed in aggregations during the mating season (Nakashima 1995). Therefore it is likely that this trail-following behaviour may also be involved in mate searching in P. scabra based on observations made during the present study and this would be a interesting area for future study.
|