Respiratory System
Aquatic hermit crabs such as the diogenids, which includes Dardanus megistos, respire by exchanging oxygen and waste carbon dioxide across gills, much like fishes and many other aquatic invertebrates. Like the gills of these other organisms, or indeed our own lungs, surface area is maximised to meet respiratory demands and circulating blood transports gases to and from the respiratory structure (Farrelly and Greenaway 2005). The gills of hermit crabs and most other organisms are not immediately obvious, else being vulnerable to attack by predators. In hermit crabs the gills are concealed in chambers below a region of the protective carapace of the cephalothorax known as the branchiostegite (Ruppert et al. 2004). The additional protection of the gastropod shell allows the branchiostegites of hermit crabs to be less calcified than those of other decapods, with heavier calcification towards the anterior (Farrelly and Greenaway 2005, Saxena 2005).

|
|
The gills of D. megistos are phyllobranchiate in form. This arthrobranch originates as an epipod, is spindle shaped and lamellate, with blood supply through the axis. The lamellae themselves are also well vascularised. Illustration: Storm Martin 2012 with reference to Tudge et al. 2004 and Saxena 2005
|
All diogenids have ten arthrobranchs (Poore 2004). Arthrobranch gills are attached to the articular membrane (Poore 2004), the soft flexible cuticle hidden beneath the protective carapace (Ruppert et al. 2004). In diogenids two pairs of arthrobranchs are found on each of five thoracic segments, specifically somites 3-7, associated with the third maxilliped through to the fourth pereopod (Poore 2004). Most diogenids, including those of Dardanus, also have a further three pleurobranchs, attached to the side of the thoracic cuticle (pleurite) of somites 5-7, carrying pereopods 2-4 (Poore 2004). Therefore, in total, D. megistos has 26 gills, 13 per side. All these gills in fact originate as epipods; projections arising from the coxa (basal segment) of the appendages (Ruppert et al. 2004). During embryogenesis these migrate to their final positions, either the pleurites or the articular membrane (Ruppert et al. 2004).
Because these gills originate as envaginations of the appendage cuticle, they are in fact cuticularised themselves (Ruppert et al. 2004). While the most obvious parts of the arthropod exoskeleton are hard and inflexible, the cuticle of the gills is extremely thin, allowing the permeability to gases required for respiration (Ruppert et al. 2004). The gills of diogenid hermit crabs have a form described as phyllobranchiate (McLaughlin et al. 2007). Each gill has an axis from which the paired gill filaments, or lamellae, extend (Ruppert et al. 2004, Farrelly and Greenaway 2005). The lamellae diminish in size towards both ends of the axis, giving a spindle form (Farrelly and Greenaway 2005). Blood channels run the length of the axis, transporting oxygen and carbon dioxide from and to the gills for respiration (Ruppert et al. 2004) and the circulatory vessels of the lamellae themselves are well developed into an extensive network (Farrelly and Greenaway 2005). Afferent branchial vessels carry blood to the gills from the infrabranchial sinuses and after exchanging of gases across the gill cuticle, blood is transported via efferent vessels to the pericardial sinus, the sinus associated with the heart which then circulates the oxygenated blood around the body (Farrelly and Greenaway 2005). The respiratory pigment of decapod crustaceans is not haemoglobin as in ourselves, but haemocyanin (Saxena 2005).

|
|
When extended from the shell, the branchiostegites, or gill chambers, become obvious as lateral bulges of the thoracic carapace. Photo: Storm Martin, Heron Island, 2012
|
While housing the gills within the gill chambers protects them from predators it has the consequences of limiting water flow across the respiratory surface, offering an appealing home to epibionts and parasites (Bauer 1981) and being susceptible to filling with sediment due to a benthic lifecycle (Saxena 2005). The arrangement of the gills establishes a continuous curtain spanning the branchial chamber, separating it into two spaces, known as the hypobranchial and epibranchial chambers (Ruppert 2004, Saxena 2005). Water is drawn through inhalant apertures of the carapace into the hypobranchial chamber, across the gills into the epibranchial chamber and then leaves via the exhalant apertures (Ruppert 2004, Saxena 2005). The inhalant apertures are located at the base of the pereopods and the two exhalant apertures are located anteriorly on the buccal frame associated with the mouth (Saxena 2005). The current drawing water through the gill chambers is established by the rapid beating (Saxena 2005) of the scaphognathite, a paddle-like epipod of the maxillae which resides in the anterior of the exhalant (epibranchial) chamber (Ruppert 2004).
The inhalant apertures are lined by a filter of setae, excluding sediment and allowing burrowing (Ruppert 2004). However, very fine filtering is not possible without impeding water flow (Bauer 1981) and so aperture setae are not exclusively relied upon for preventing sediment build-up. To remove sediment and also epibionts or parasites, decapods periodically reverse the respiratory current, flushing the gill chamber (Bauer 1981). The gills and the scaphognathites are also regularly groomed. The scaphognathites are wiped with the third maxillipeds, in the same motion which also cleans the antennules (Thatje et al. 2010). The gills are groomed by the last (fifth) pereopod, which in hermit crabs has become specialised for this purpose, is reduced and is often held within the gill chamber (Bauer 1981).
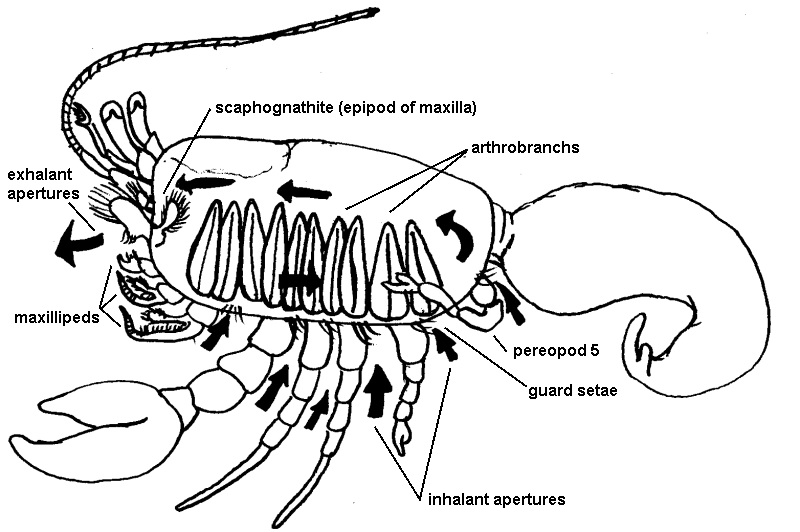
|
|
The respiratoy system overview. The beating scaphognathite draws water in through setae guarded apertures of the pereopod attachment site to the body. The water circulates across the curtain of gills and exits via apertures of the buccal frame. Flow can be reversed for flushing clean. Only arthrobranchs are depicted here. Illustration: Storm Martin 2012 with reference to Tudge et al. 2004 and Saxena 2005
|
|