|
|
|
Littoraria (Littorinopsis) filosa, the Thin Mangrove Periwinkle
|
|
Gustavo Zoppello Toffoli 2015
|
|
|
|
Summary | |
The periwinkle term refers to species of sea snails
that are allocated in the family Littorinidae, class Gastropoda. They usually
occupy intertidal ambient and can be found in rocky, sand, mud and mangrove
substrata all around the globe. Due to the characteristics of the intertidal
zone, these animals have specific adaptations to deal with salinity and
temperature variations, desiccation, wave action and predators. These characteristic
makes periwinkles a good object of study, therefore ecology, physiology and
morphology studies of periwinkles are well developed as well as the cladistics
and evolution study in order to promote the understanding of the success of
these worldwide spread animals.
The thin periwinkle Littoraria (Littorinopsis) filosa is endemic from the east coast of
Australia and is known for its polymorphic shells which can varies from yellow
to pink. Their shell can achieve no more than 3 cm height and they are usually
found in mangrove leaves where they feed. The specie prefer to be on high
heights far from the water and just go down to release larvae.
Due to the mangrove association, these gastropods
can be used as indicative factors of the mangrove characteristics what can be a
useful tool for the study of the impact caused by ocean pollution and
destruction of mangroves.
 Photo: Thin periwinkle (Littoraria filosa). Gustavo Zoppello Toffoli, Wellington Point - Queensland,
2015
Photo: Thin periwinkle (Littoraria filosa). Gustavo Zoppello Toffoli, Wellington Point - Queensland,
2015
|
|
|
Physical Description |
Body Description | |
The
bases of the physical description of periwinkles are often related with the
spiral univalve shell shape. As gastropods, they can withdraw they body inside
the calcareous shell produced by glands in the mantle. The gastropod’s shells
have a specific shape with can be used for identification and cladistics
studies (BRUSCA & BRUSCA, 2002). The Littorinidae family is known for own
large, thin shells with variable coloration and sometimes brightly (REID,
1986).
The
body has bilateral symmetry and is divided in head, visceral mass (inside the
shell) and foot used of locomotion, Figure
1, which is the most easy viewed soft part of a gastropod (RUPPERT et al., 2004).
 Photo: L. filosa lateral view. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
Photo: L. filosa lateral view. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
Body Morphology | |
The L. filosa
basic bauplan as well as a gastropod, is defined for an anterior part, the
head, the visceral mass that stays inside the shell and a big muscular foot,
used in the movement of these animals (BRUSCA & BRUSCA, 2002).
Sclerotized cuticle, single layer epidermis and
muscles are the three layers of the body of a gastropod. The water loss is a
constant problem for these animals as they do not have a specialized cuticle to
avoid it but they may withdraw inside their shells to prevent water loss.
Terrestrial species also have to spend with mucus that facilitate the water
wasted (BRUSCA & BRUSCA, 2002). The dorsal body is surrounded by the mantle,
where are located the shell glands, responsible for the shell secretion. There
is an invagination of the visceral mass that forms with the shell, the mantle cavity,
where is located the gills, the anus, nephridiopores and gonopores (RUPPERT et al., 2004).
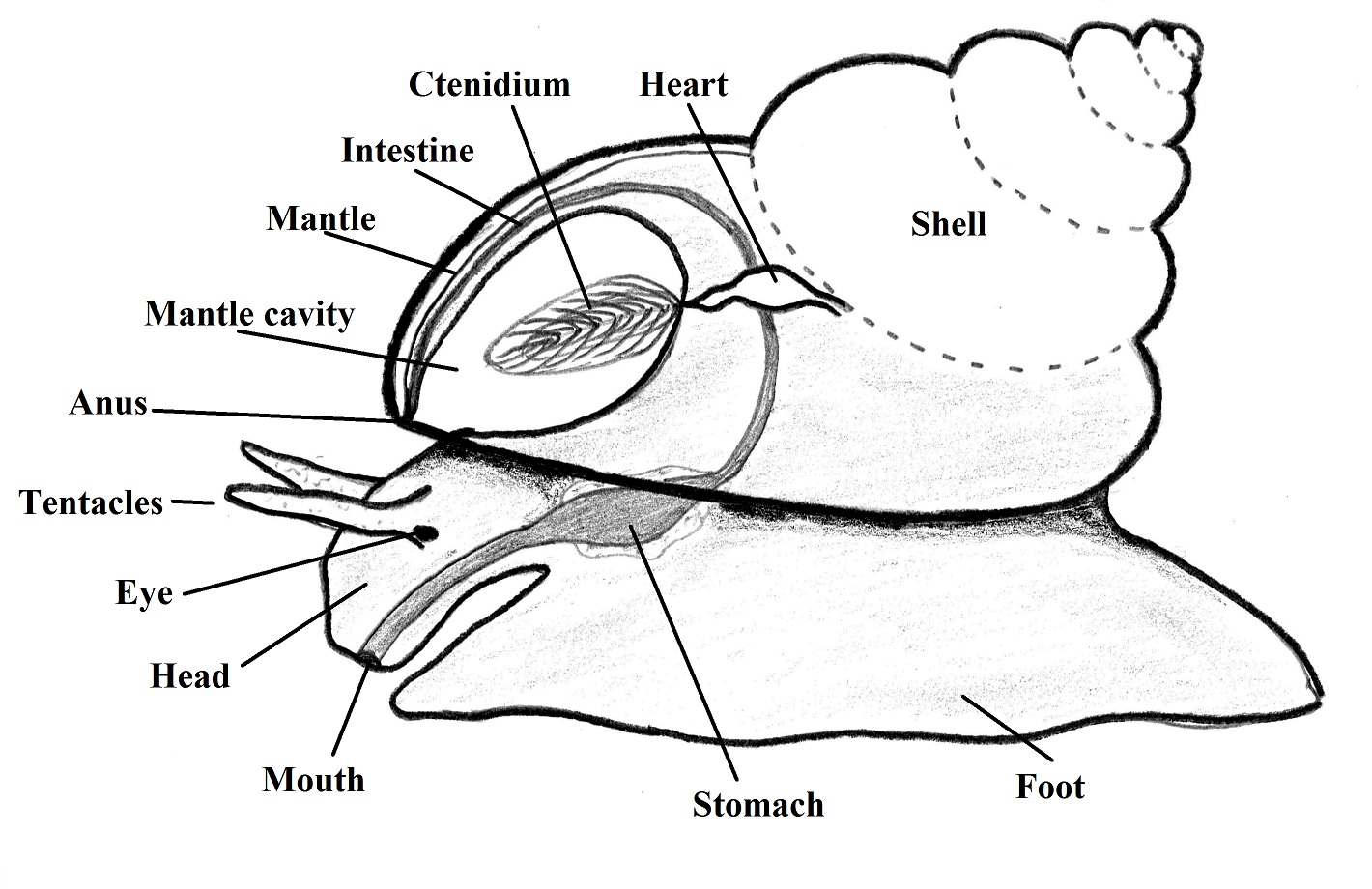
Figure
1. Representation of the gastropod basic bauplan. Illustration by Gustavo
Zoppello Toffoli, 2015 with reference to (BRUSCA & BRUSCA, 2002).
The color of the head and the foot of the gastropods
are used for taxonomical propose too. In Littoraria
species, the color of the body are linked with the polymorphic color of the
shells, as described by Reid (1986):
“For example, in Littoraria filosa the animals with pure
yellow shells are entirely unpigmented, in darker shells animal pigmentation
becomes more pronounced, especially on the head, until animals are dark grey in
brown shells”
In L. filosa the
peculiar feeding structure of the gastropods is pink colored and is called
radula, located in the head as well as the sensory structures that composed by
a pair of eyes in the base of a pair of tentacles Figure 1 (REID, 1986). The operculum is the structure used for
close the animal inside of the shell when it is windrowed. This structure is
used for prevent desiccation and as a shield against predators. The operculum in
L. filosa is thin and paucispiral and
very similar to the other species from the genus Littoraria (REID, 1986).
 Photo: Frontal view of a L. filosa, is possible to see part of the head with two tactile tentacles; the eye (left side); part of the buccal mass and the frontal edge of the foot. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
Photo: Frontal view of a L. filosa, is possible to see part of the head with two tactile tentacles; the eye (left side); part of the buccal mass and the frontal edge of the foot. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
Shell Morphology | |
Shell
Morphology
Gastropods are univalve animals and their
shells can vary in size from microscopic to more than 40 cm in some species. The
distinctive characteristic of the group is the conical spiral around the axis
called columella that forms the spire divided in whorls by the sutures that are
the lines where the whorls attach to the others (BRUSCA & BRUSCA, 2002).
The body whorl is in the base of the shell and is where the aperture is located
which is used for body retraction or extension by action of the columellar
muscle (RUPPERT et al., 2004). See Figure 1:
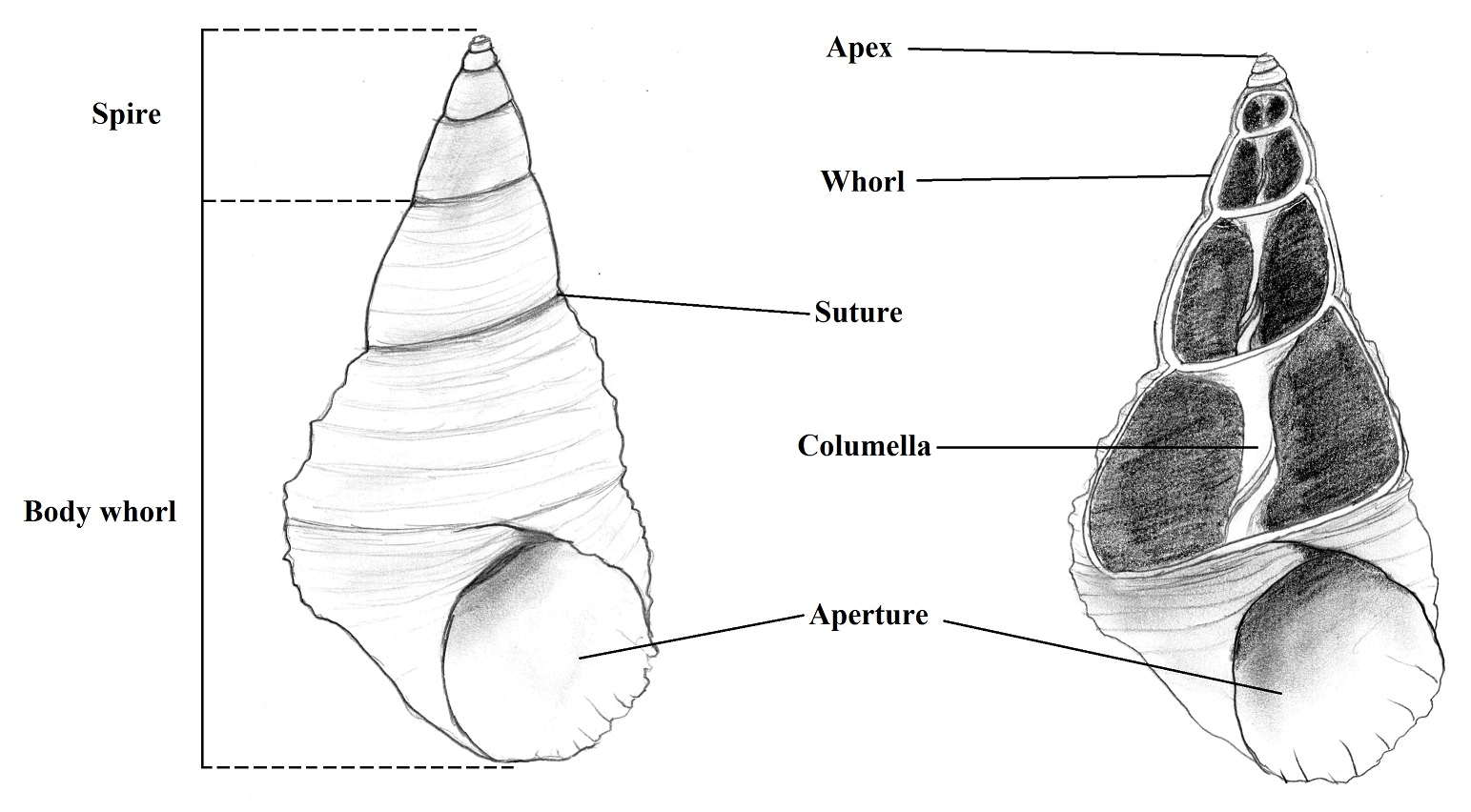 Figure 1. Dextral shell of L. filosa. (left - Intact shell; right - longitudinal section). Illustration by Gustavo Zoppello Toffoli, 2015, modified of Ruppert et al. 2004.
Figure 1. Dextral shell of L. filosa. (left - Intact shell; right - longitudinal section). Illustration by Gustavo Zoppello Toffoli, 2015, modified of Ruppert et al. 2004.
The shell in periwinkles are constituted from
outside to inside by a thin organic periostracum, two or three prismatic layers
that differs in their composition but are all caucareous and the nacreous layer
made of calcareous and conchin lamellas as the Figure 2
shows (BRUSCA & BRUSCA, 2002).
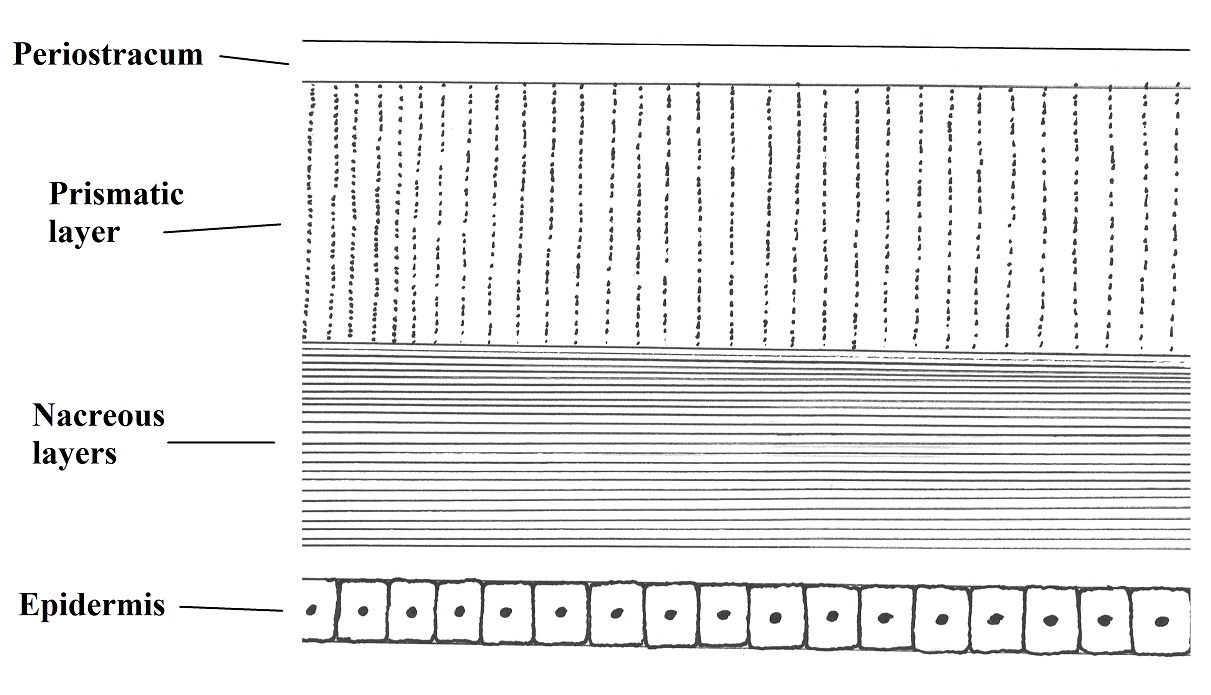 Figure 2. Common constitution of a gastropod shell (section). Illustred by Gustavo
Figure 2. Common constitution of a gastropod shell (section). Illustred by Gustavo
Zoppello Toffoli, 2015 based on Brusca & Brusca, 2002)
Is important to remember that the shell is produced
by addiction of organic or inorganic elements and the different material
deposition in the inner and in the outer layers is responsible for the spiral
formation and it is metabolic expansive requiring substantial sources of the ambient
(RUPPERT et al., 2004). Although for
example, calcium carbonate is common abundant in mangrove environment even in
high tidal levels (REID, 1986)
 Photo: Shell deposition, is possible to see a transparent part which correspond to the formatio of a new whorl. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
Photo: Shell deposition, is possible to see a transparent part which correspond to the formatio of a new whorl. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
The color varies according to the periostracum
pigments or by pigments in the caucareous layers. The origin of the pigments
may be related with the diet or may be synthetized by the gastropod. This
pigment addiction can be related with cryptic behaviors (RUPPERT et al., 2004).
In Littoraria
genus are mainly associated with mangrove habitats, the shape of the shells are
apparently uniform despite some variations, and there are some evidences that
the planktonic dispersal behavior is responsible for this combined
characteristic. L. filosa has a thin,
dextral shell (clockwise spiraled).
As a supralittoral specie, L. filosa has a thin, light shell, which might be for decrease the
risk of dislodgment, as they stay attached in the leaves just by the action of
the mucus. Another hypothesis about the thickness of the shell may be related
with predators in low levels which selecting individuals for thicker shells,
letting the individuals with less thicker shells occupy high levels (REID,
1986). The character of a supralittoral specie that occupy high leaves may be
influencing in the size of the shell. Without wave action, the shells can be
bigger and will not be washed out from the leaves, also, the higher tolerance
of larger individuals against physical conditions as suggested by Vermeij,
1973b in Reid (1986). The more narrowed shape of the shell may be influenced by
an allometric result of the increase of the size (REID, 1986).
Works with L.
filosa has shown that in breeding periods, the shell growth ceases, the
apertural lip is flared and thickened, and after the growth continued, it is
possible to be seen in some adult individuals (REID, 1986). However, this
flared characteristic is not seen in all adult individuals what may be,
according to Sewel (1924) cited by Reid, 1986, related to the habit of
occupying high tidal levels as the breeding period occurs at the same time of
high rainfalls and temperatures what may interfere in the growing rates. Then,
the flaring and thickening of the outer lip of the aperture may not be related
with breeding periods.
The sexual dimorphism is not very apparent and is
variable. Despite the fact that females have apparently larger shells, and
males have lower spire height (REID, 1986).
Despite the not useful characteristics of the
protoconch (first whorl) that is smooth and from the second whorl (protoconch
2) for taxonomy, is important to remember that the protoconch is secreted by
planktonic larvae and due to the protection of the mangrove habitat, this first
whorl can be found even in adult individuals (REID, 1986).
 Photo: Visualization of the protoconch (the first whorl) in both individuals. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015 Photo: Visualization of the protoconch (the first whorl) in both individuals. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
The shell sculpture in Littoraria is the most useful and important taxonomic
characteristic, used for distinguish the diverse species as is possible to see
in the diagnosis part. However, some species cannot be distinguished by shell
characteristics needing the verification of the geographical range (REID,
1986). Due to the high intertidal preferred position in outer mangrove trees,
the control of the body temperature in L.
filosa is very important. Its shell is strong sculpted, a characteristic
that can improve the loss of temperature by increasing the superficial area
(REID, 1986).
L.filosa
lives on the high foliage of mangroves trees and is considered one of the most
polymorphic Littoraria species. The
different colour patterns can be found on the same tree. In Avicennia marina (the preferred tree of L. filosa), the color varies from light ground color, yellow and orange pink which
defines the variation spectrum of the colors (REID, 1986).
Many studies about the polymorphism of the shell
color are already done and some thermal regulations evidences are found by Reid
(1987) and Parsonage & Hughes (2001). Despite the suppositions relating
this characteristic with crypsis selection by birds (JOHANNESSON &
EKENDAHL, 2001) and at lower levels of the trunks by crabs (REID, 1986), the
main factor ruling the polymorphism in L.
filosa is the parasitoid fly Sarcophaga
megafilosia as reported by (MCKILLUP & MCKILLUP, 2002 and 2007).

Photo: Some examples of shell polymorphism in L. filosa. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
Diagnosis and Key to Shells | |
Diagnosis
The diagnosis of Littoraria
filosa as postulated by Reid, 1986:
"Shell: thin; columella narrow, rounded; primary grooves 5-7(9), spacing
markedly unequal, increasing anteriorly; last whorl with 9-11 strong, narrow
carinae, the spaces between with numerous irregularly spaced spiral grooves;
colour highly polymorphic, brown, yellow or orange pink, with or without
pattern of brown dashes aligned at suture to form short stripes, numbering
10-12 on last whorl. Animal: penis bifurcate, filament large, narrowing only at
tip; ovoviviparous."
Key to Shells
According to the
key for shell identification proposed by Reid, 1986 L. filosa is identified by the following charachteristics:
1 Columella
narrow, rounded, not excavated . . . . . 2
- Columella
excavated or flattened, usually wide . . . . 8
2 Sculpture
of 9-11 narrow carinae on last whorl; colour polymorphic L. filosa
- Sculpture of low
or rounded ribs, or numerous fine riblets
. . . 3
 Photo: Thin periwinkle (Littoraria filosa). Gustavo Zoppello Toffoli, Wellington Point, Queensland,
2015
Photo: Thin periwinkle (Littoraria filosa). Gustavo Zoppello Toffoli, Wellington Point, Queensland,
2015
|
|
|
|
Ecology |
Distribution | |
Adapted to a terrestrial life, L. filosa has as its preferred substrate,
the leaves of the grey mangrove Avicennia
marina where the group achieve the greatest diversity and abundance (REID,
1986). However, it shares the habitat with other littorinids, each specie
specialized for a specific height level related with the tide inundation and
rain that have been showed as the most important factors influencing the
zonation of these periwinkles. (REID, 1986).
The pattern of distribution through
the mangrove tree species appears to be related with the preferred height, some
leaf dwelling species as L. filosa, L.
luteola and L. albicans are
described as more frequent on the leaves of A.
marina whereas some other species that prefer to live on the trunks or on
the roots are more dispersed (REID, 1986). In relation to the horizontal
zonation a pattern has been noticed by the authors that Littorinids occurs in
all the mangrove forest but they begin to be scarce far away from the sea edge
(REID, 1986) and L. filosa is less
abundant in more wave-exposed mangrove areas (REID, 1985).

Photo: L. filosa can be found in various locations in a tree. They can be found alone or in patches. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
Predators | |
The main tree periwinkles’
predators are birds and crabs. The crab predation decrease with the height,
then, for the high level tree L. filosa, this
kind of predation is just important when the females are releasing the larvaes
and in dry conditions when the periwinkles goes down to obtain moisture. The
birds may cause more damage as they usually search for food flying. However,
the L. filosa use to graze on the
underside of the leaves what difficult the detection for the flying predators.
 Photo: Representat of the genera Sarcophaga which the parasitoid fly is part of. Gustavo Zoppello
Photo: Representat of the genera Sarcophaga which the parasitoid fly is part of. Gustavo Zoppello
Toffoli, Wellington Point - Queensland, 2015.
Researches have been shown that the
principal predator of L. filosa is
the parasitoid fly Sarcophaga megafilosia
as reported by (MCKILLUP & MCKILLUP, 2002 and 2007). The polymorphic
characteristic of the shell have been used for avoid the deposition of larvaes
near from the snail by the female flies that will occasionally kill the
periwinkle (MCKILLUP & MCKILLUP, 2002)
 Photo: Example of how the crypsis effect work: A yellow shell periwinkle on a yellow leaf. Gustavo Zoppello Toffoli, Tinchi Tamba Wetlands - Queensland, 2015
Photo: Example of how the crypsis effect work: A yellow shell periwinkle on a yellow leaf. Gustavo Zoppello Toffoli, Tinchi Tamba Wetlands - Queensland, 2015
|
|
|
|
Life History and Behaviour |
Habitat and Feeding | |
From
the Littorinidae family, periwinkles
are found in the intertidal habitat as rocky shores, mangroves and saltmarshes
all around the world. L. filosa is a high-level
tree-obligate periwinkle adapted to a terrestrial life able to live up to 3
meters above ground (Avicennia zone,
Magnetic Island, Queensland) that climbs to avoid submersion during the high
tides (REID, 1986). Mostly found on Avicennia
marina (grey mangrove) leaves (Photo) and sometimes trunk
(REID, 1986; RUPPERT et al., 2004).

Photo: A. marina mangrove habitat. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
The snails stays in high levels even in low tides
just descending to the water to get moisture or in order to release the
larvaes. As an adaptation to the semi-terrestrial habit, L. filosa developed an ovoviviparous characteristic. The female
maintain the embryos between the gills and when they are developed, she goes
down to the water and release the veliger larvaes. The horizontal mobility is
restricted to trees with branches in contact due to the inability of these
periwinkles to move on mud as described by Reid (1986).
Depending on the weather conditions, periwinkles may
face some dry conditions, which makes them withdraw inside the shell and
staying attached to the leaves by action of the mucus (REID, 1986), while the
entire animal is inside the shell, closed by the operculum.
The L. filosa is
a leaf dwelling specie that feed on the underside of A. marina leaves, scraping the leaf hairs seldom opening holes
through the leaves (REID, 1986). For some species that lives on other mangrove
tree species, the hairs from underside of A.
marina leaves may not be the main food source, however, L. filosa practically is not found
grazing on other mangrove tree species, then, probably the A. marina leaves hair is probably an important source of food for
this specie.
 Photo: L. filosa can grazing on the underside of a A. marina leaf. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
Photo: L. filosa can grazing on the underside of a A. marina leaf. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
Copulation | |
As described by many authors before Reid (1986), the
copulation only occurs in moisture conditions. The behavior of L. filosa is the same as described of
other Littorinids where the males look for females attaching to any other
individual as they are only able to verify the sex by trying to insert the
penis into the bursa cupulatrix.
Intraspecific mating occurs without evidences of
hybrids individulas. In the case of a meeting of males, they rapidly separates.
The male lean to the anterior right side of the female body and inserts the
penis into the aperture, the act is short but the male remains attached for
several hours (REID, 1986).
|
|
|
|
Anatomy and Physiology |
The Body Features | |
The Body Features
As gastropods, L.
filosa are coelomate animals, but in their body, the coelom can just be
found as a vestige structure surround the heart and part of the intestine as
the pericardial chamber and the perivisceral coelom respectively whereas the
main chamber consist of several cavities composing an open circulatory system
(BRUSCA & BRUSCA, 2002).
An important feature of the periwinkle
bauplan is the planispiral coiling of their body inside the shell, forming
whorls that are positioned outside of the antecessor whorl. Although no live
gastropod has a planispiral shel, instead of that, their shell varies in more
than one plan of an asymmetric whorl formation (RUPPERT et
al., 2004).
These animals must deal with a heavy
structure attached in their dorsum, the shell. Due to the planispiral shape,
the center of gravity of the body is taken upwards what makes difficult to
maintain the shell erect (RUPPERT et al.,
2004). The asymmetrical conispiral shape of the shell solved this problem.
Instead the shell grow upwards, it grows dislocated from the body axis,
repositioning the center of gravity (Figure 1) and facilitating
the movement of the animal (RUPPERT et al.,
2004).
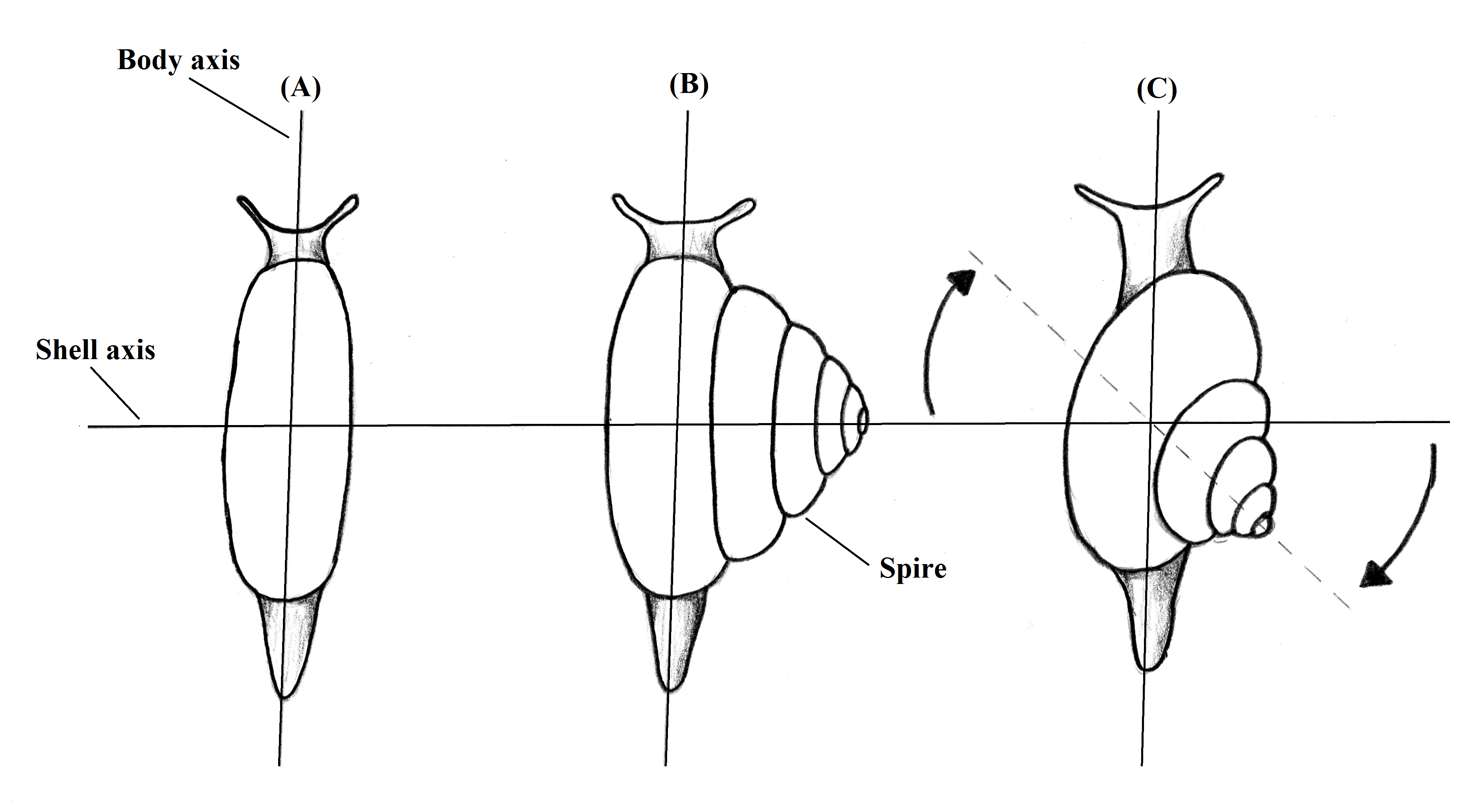
Figure 1. Representation of the evolution of the asymmetry in gastropod shells. Vision from above. Illustration by Gustavo Zoppello Toffoli, 2015 based on Ruppert et al. 2004.
A notable step in the gastropod
development that is also one of the synapomorphies of the modern groups is the
phenomenon called torsion. A remarkable twist of 180° counterclockwise
(that occurs during the late veliger larval development without change the head
and foot positions. The visceral mass is twisted changing the right/left side
of structures when compared the larvae with the adult. The torsion of the gut
let it with a U-shape and the nerves are also twisted. In the end of the
process the gills, nephriopores, hypobranchial glands, osphradia and the anus
are now posterior positioned as is possible to see in the Figure 2 (RUPPERT
et al., 2004; BRUSCA & BRUSCA, 2002).
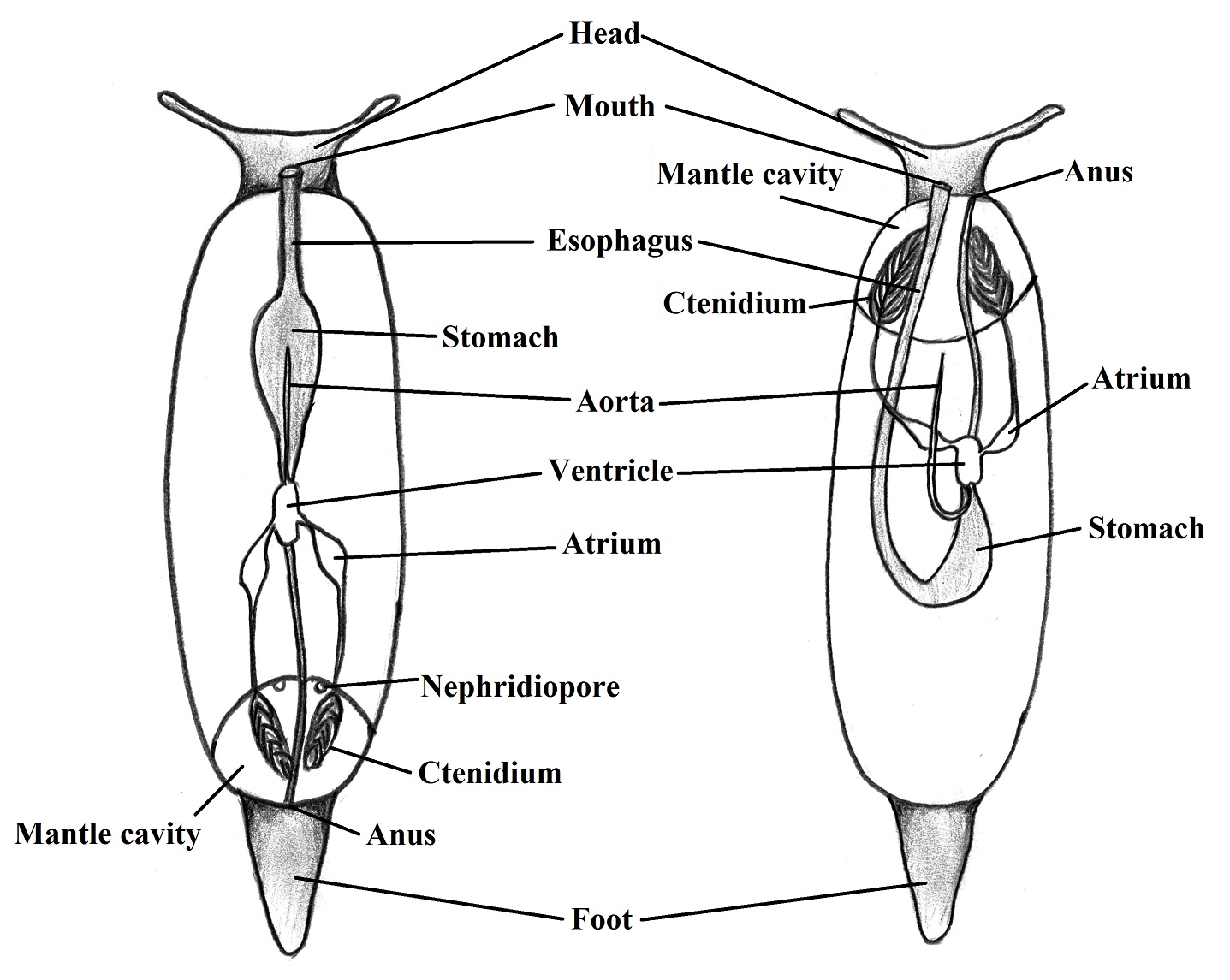
Figure 2. Causes of the torsion in gastropods body. (left - hypothetical untorted ancestral; right -
gastropod after torsion. Illustrated by Gustavo Zoppello Toffoli, 2015 based on Ruppert et al. 2004.
Some theories have been used to
justify this torsion as that the posterior position of the mantle cavity may
improve ventilation, the posterior position of the osphradium allow the animal
to verify the ambient before rather than after entering in the water. However,
the twist, put the anus and nephriopores in a position close to the mouth and
head what caused a sanitation issue that interfere with the gills/lung
ventilation, some specializations evolved to deal with this problem (RUPPERT
et al., 2004).
|
|
|
Locomotion | |
Foot
The foot is a
muscular structure responsible for the movement of the gastropods that can be
related with feeding, prey capture, reproduction and in females, molding of the
eggs. It is linked by the columellar muscle that is responsible for extension
and retraction of the foot. In the sole, there are the tarsos muscles directly responsible
for the locomotion (RUPPERT et al., 2004).

Photo: The bottom part of a L. filosa, is possible to see the large muscular foot. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
Movement
The sole is broad and flat, properly
for movement in diverse substrates. There are glands that secretes mucus that
facilitate the attachment and locomotion The snail can chemically change
consistence of the mucus from sticky gel to liquid depending on the need (RUPPERT et al.,
2004). The L. filosa as a tree living
snail, is considered a hard bottom terrestrial gastropod that uses waves of the
tarsos muscles to move on the leaves and trunks (RUPPERT et al., 2004). The direction of the
movement of the sole muscles varies depending on the species, it can be in
direct waves, retrograde waves or bipedal when the left and the right side of
the foot alternate the direction of the waves (BRUSCA & BRUSCA, 2002).
Video: Movement of a L. filosa on a leaf. Notice the amount of mucus and the tactile function of the tentacles. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
Feeding and Digestion | |
Radula
Radula is the unique mollusk structure used for
feeding. Is constituted by a band of recurved chitinous teeth. Associated with
strong muscles, the radula projects from the pharynx (BRUSCA & BRUSCA, 2002)
and is used for L. filosa snails scrape
the surfaces of the leaves where they feed (REID, 1986). The radula is an
important feature for cladistics and may be the characteristic that supports
some groups (BRUSCA & BRUSCA, 2002).
The description of the L. filosa radula is wrote as “length to 17 mm, saw-toothed type,
central rachidian cusp elongate and pointed, cusps of paired teeth equilaterally
triangular and the lateral with slight gap aterior to main cusp”, by Reid
(1986).
 Photo: The radula is extended with the buccal mass in order to achieve the substrate, the red colour is due to the presence of myoglobin in the muscles. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
Photo: The radula is extended with the buccal mass in order to achieve the substrate, the red colour is due to the presence of myoglobin in the muscles. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
Gut
Periwinkles have complete guts and their digestion
is mainly extracellular. The mouth is located inside of a buccal cavity where
the radula apparatus leads. Some glands releases saliva and enzymes into the
anterior gut. The esophagus connect the foregut to the stomach that will end in
the digestive ceca where the absorption and the intracellular digestion occurs.
The intestine links the stomach region to the anus located in the mantle cavity
(BRUSCA & BRUSCA, 2002; RUPPERT et al.,
2004). See Figure 1:

Figure 1. Digestive system of a land snail. Illustred by Gustavo Zoppello Toffoli, 2015 modified of Brusca & Brusca, 2002.

Photo: Faeces of L. filosa indicated by the narrows. (fresh in the right image and dry in the left image). Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
Excretion and Osmoregulation | |
As L. filosa is
more terrestrial but still an amphibious species, their nitrogenate excreta may
be mainly ureotelic instead of ammonotelic. The kidneys of the gastropods
consists in tubular metanephrids like the annelids as Figure 1 shows (BRUSCA & BRUSCA, 2002). Due to the
torsion, the nephridiopore is located in the mantle cavity near from the anus (RUPPERT
et al., 2004).
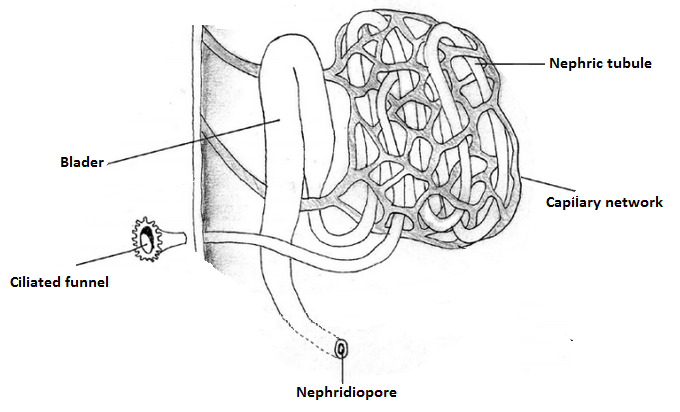
Figure 1. Metanephridium structures. Illustrated by Gustavo Zoppello Toffoli, 2012, adapted
of Hickman et al. 2004.
L.
filosa cannot avoid losing water, but in dry periods they
can decrease the metabolic rate and withdraw inside their shell and close the
operculum keeping attached to the leaf until good water source conditions (RUPPERT
et al., 2004).
|
|
|
Respiration and Circulation | |
The
gas exchange occurs through modified gills, surface of the mantle cavity and
the body surface (BRUSCA & BRUSCA, 2002). The coelom of the body of gastropods
is reduced and the principal body cavity is the hemocoel that works as an open
circulatory system which the vessels are twisted due to the torsion (RUPPERT et al., 2004). There is one muscular
atrium that receives oxygenated blood from the gills whereas the more
musculated ventricle pump it to the head and the visceral mass. The blood in gastropods
works for the internal transport with help of the hemocianin pigment but the
main role of the blood system is its function as a hydrostatic skeleton (RUPPERT
et al., 2004). The Figure 1 represents the basic circulatory system with association
to the gills.
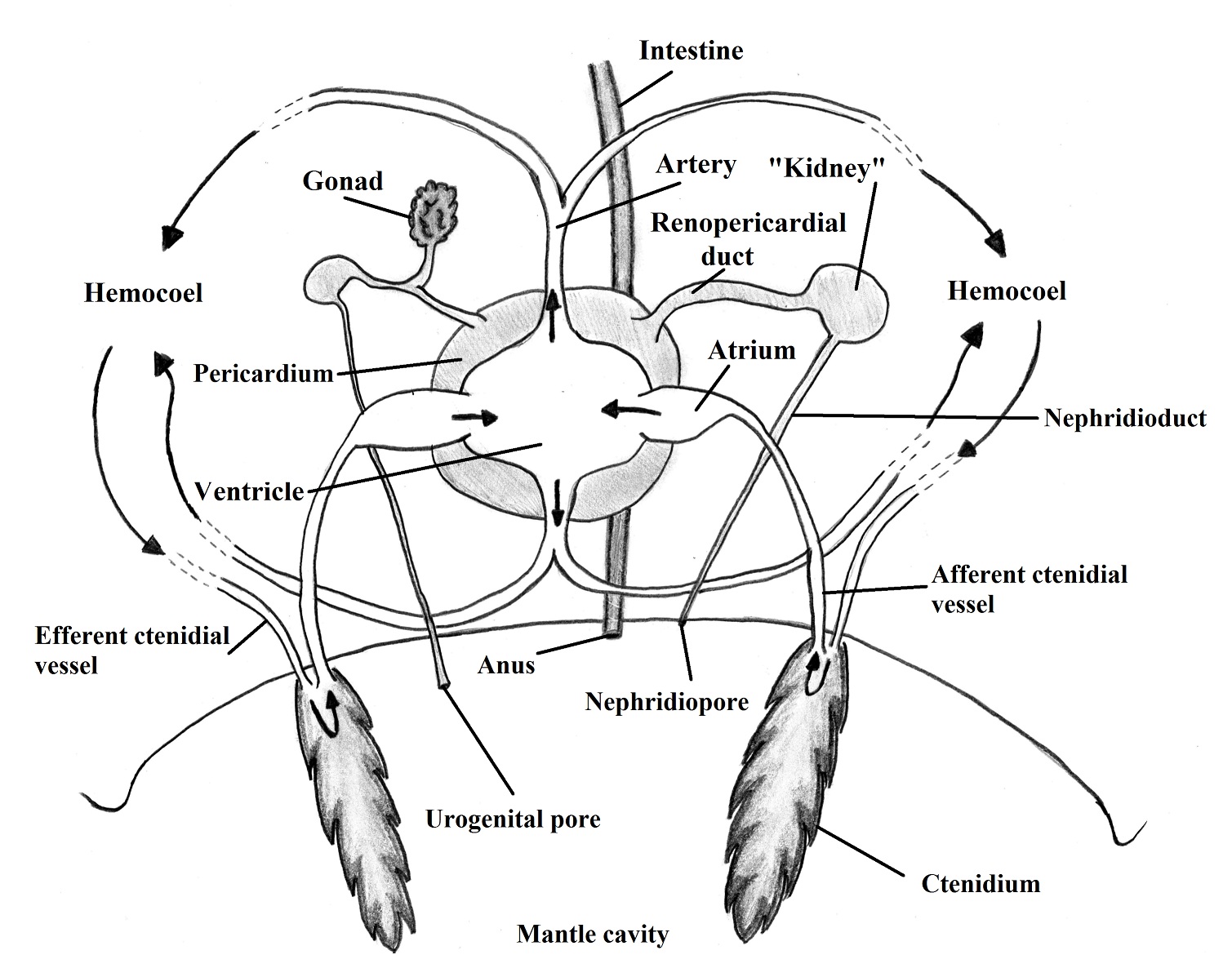
Figure 1. Circulatory system, showing the hemolymph flow. Illustred by Gustavo Zoppello Toffoli, 2015
adapted from Brusca & Brusca 2002.
|
|
|
Nervous System and Sense Organs | |
The
nervous system is composed by an anterior ganglia and paired ventral cords. Due
to the torsion, the parts of the nervous system that links the head and the
foot with the visceral mass is torted forming the visceral loop (BRUSCA &
BRUSCA, 2002). See Figure 1.
The L. filosa sense organs include a pair of simple eyes positioned in
the base of the cephalic tentacles and appear to be just for light detection, a
pair of cephalic tentacles that act as a tactile structure, osphradia that has
a sensitive chemic function related with water quality and statocystis in the
foot for movement detection (REID, 1986; RUPPERT et al., 2004).
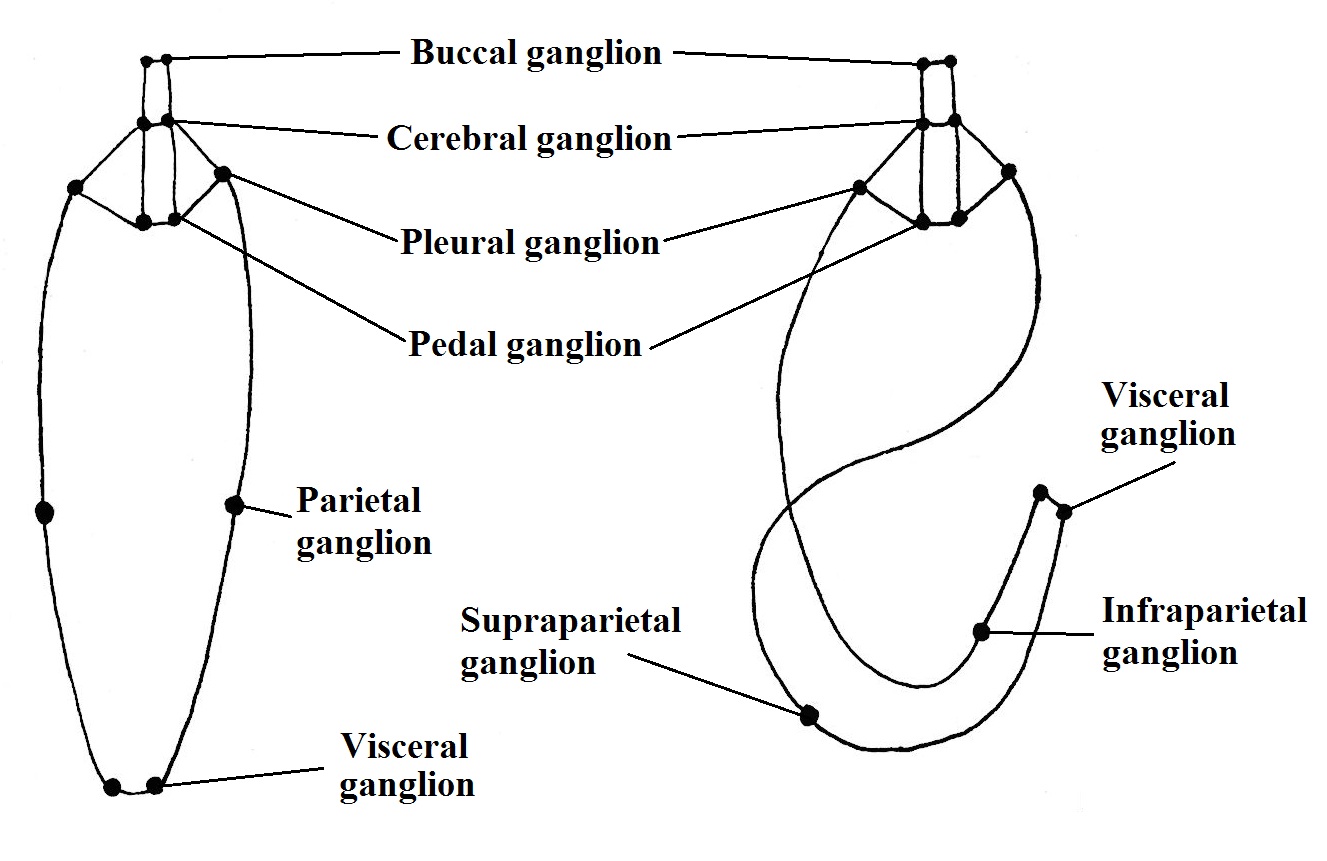
Figure 1. Ganglia disposition in hypothethical untorted ancestral (left) and torted adult of gastropod. Illustrated
by Gustavo Zoppello Toffoli, based on Brusca & Brusca 2002.
|
|
|
Reproduction | |
They
have just one gonad which is linked to the right nephridium by the gonoduct. L. filosa is a dioecious specie; the
male has a penis that stays behind to the right cephalic tentacle that gives
assistance to the sperm transference and also a prostate gland responsible for
seminal fluids production. The female has an ovary that extend through the
digestive gland, a thin, transparent oviduct and a seminal receptacle responsible
for the sperm storage and production of the egg capsule. However, as an
ovoviviparous specie, there is not a capsule covering the eggs in L. filosa.
The penis characteristic is used in
taxonomic classification as well as palial oviducts and sperm nurse cells
(REID, 1986).
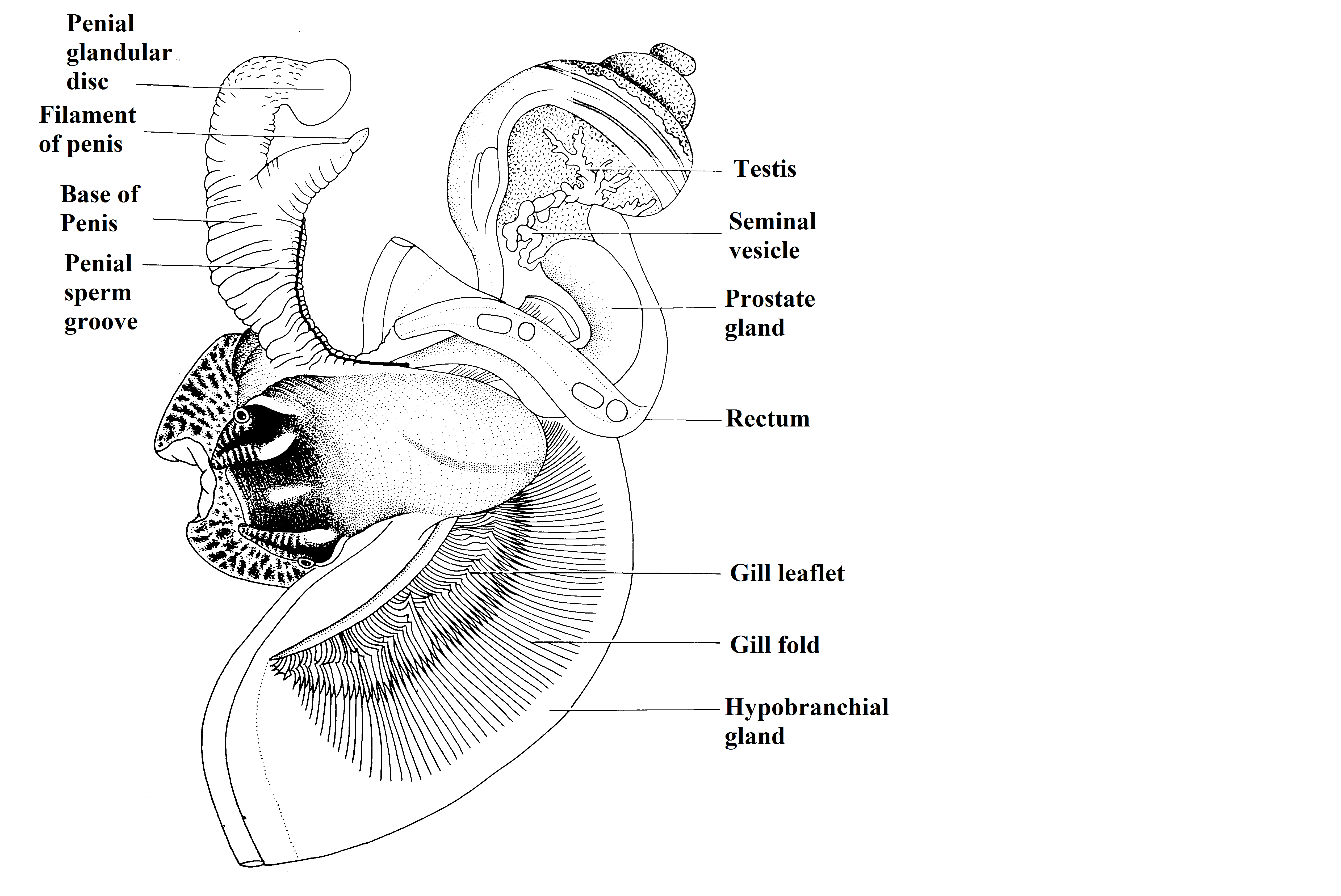
Figure 1. Male reproductive system, mantle cavity opened. Adapted from Reid, 1986
(authorized image usage).
©David G. Reid
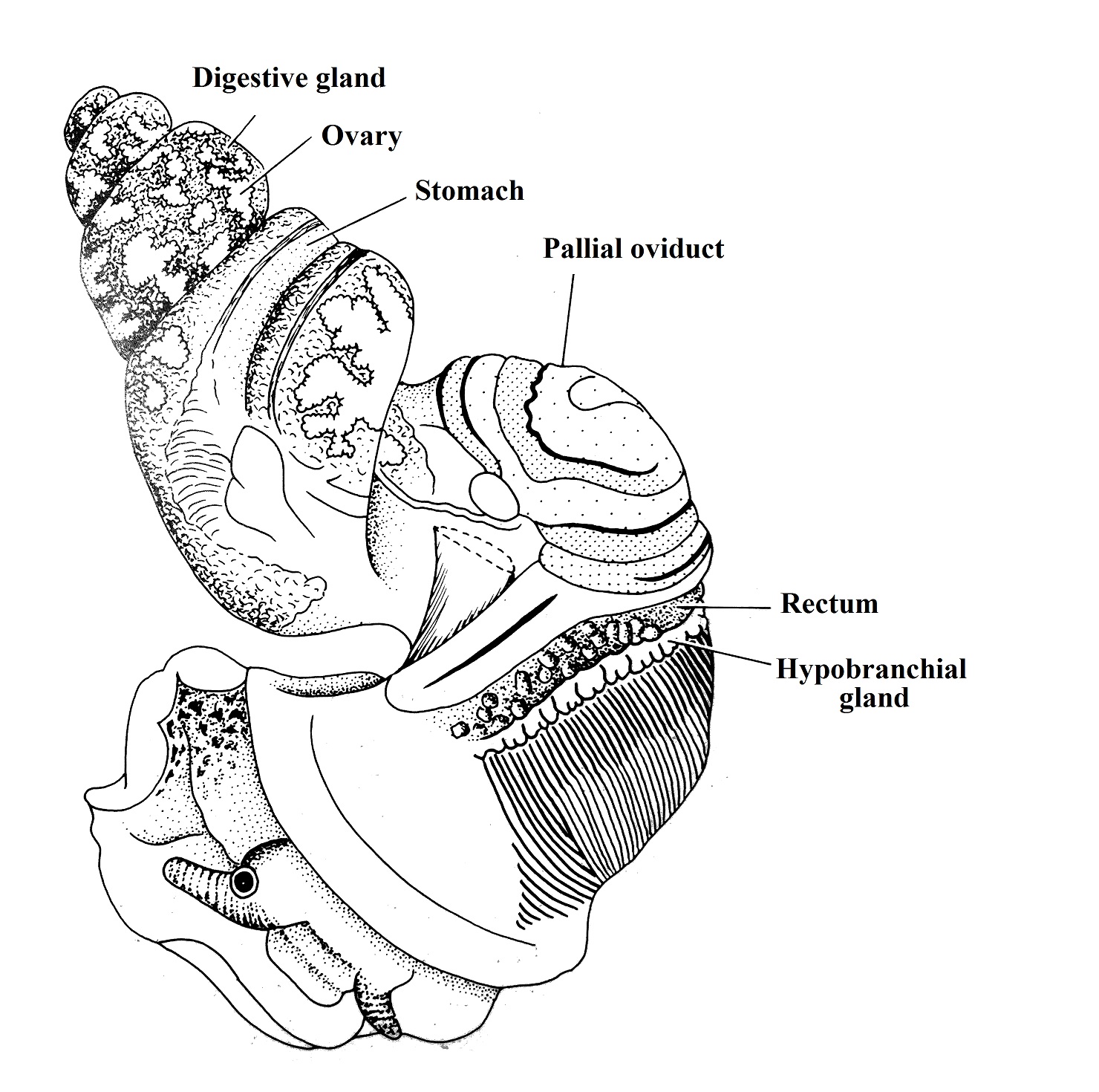
Figure 2. Female reproductive system. Animal removed from shell. Adapted from
Reid, 1986 (authorized image use). ©David G. Reid
|
|
|
Development | |
Periwinkles,
as protostomes, have a typical spiral cleavage (BRUSCA & BRUSCA, 2002). The
development in L. filosa is indirect
with formation of a larval stage called trochophore similar to the annelids,
then, giving way to a molluscan exclusive larval stage called veliger that
already has some of the adult structures (BRUSCA & BRUSCA, 2002). See Figure 1 below:
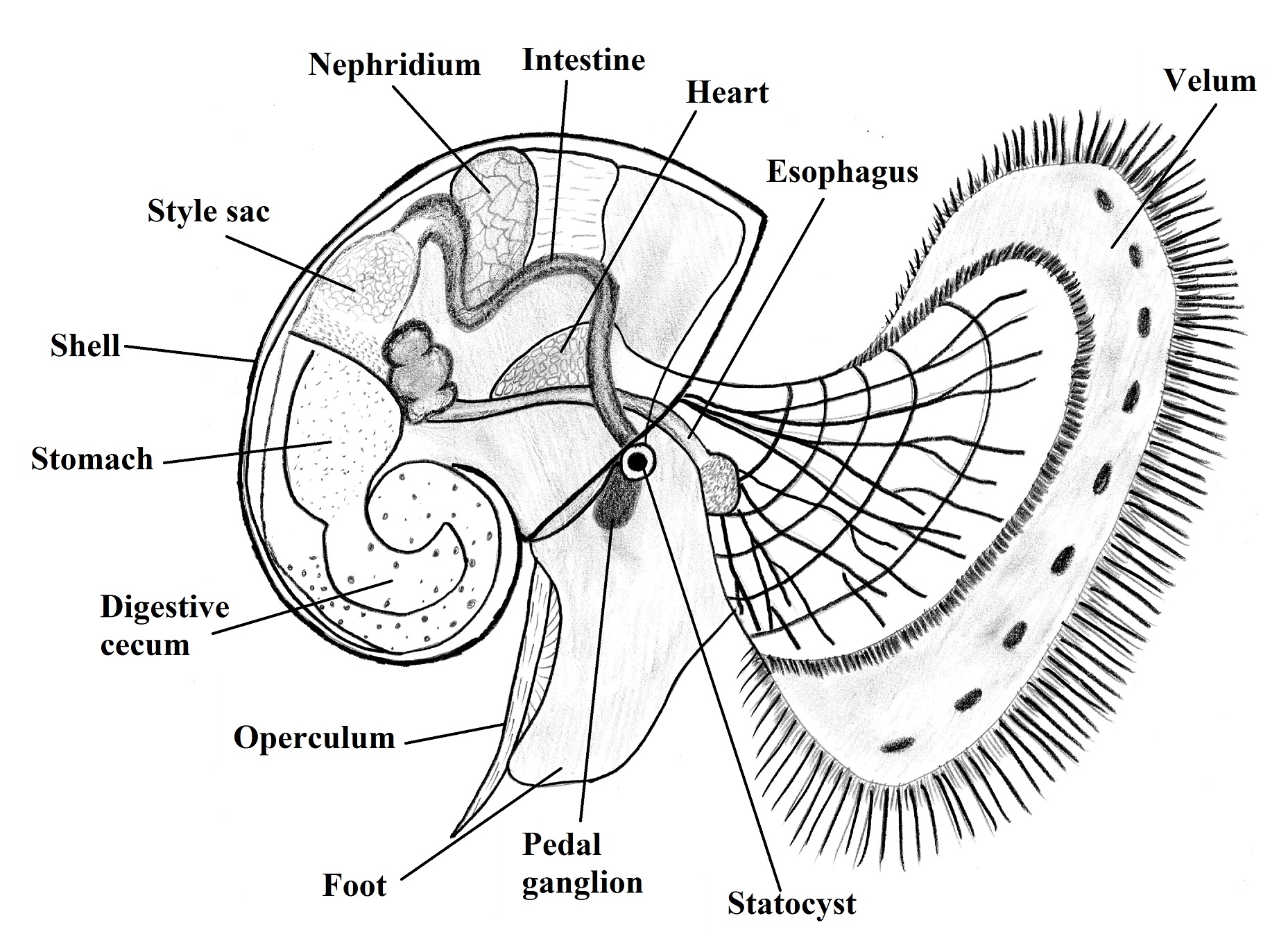
Figure 1. Model of a veliger larvae of a snail (lateral view). Illustred by Gustavo Zoppello Toffoli, based on Ruppert
et al., 2004.
As an adaptation to the semi-terrestrial habit, L. filosa developed an ovoviviparous characteristic
releasing the veliger larvaes into the water for a planktonic stage of life (REID,
1986). This type of larvae is responsible for the dispersion of the specie and
it is interfered by the ocean currents (RUPPERT et al., 2004).
The settlement process is not well studied but due
to the distribution, probably it occurs in mangrove trunks, roots and
pneumatophores by a chemical selective process. After the settlement, if
favorable conditions, the larvae will metamorphose to the adult form (RUPPERT et al., 2004).
|
|
|
|
Evolution and Systematics | |
The
evolution of mollusks is quite complex due to the high diversity and because of
polyphyletic and/or paraphyletic division of the classes. As they share the
principals protostome features for example the spiral cleavage, schizocoely and
the trochophore larva. These symplesiomorphic characteristics cannot be used
for differentiate the phylum (BRUSCA & BRUSCA, 2002).
The main characteristics that differ
mollusks and annelids are that the mollusks do not present segmentation and
present an open circulatory system as well as the modification of the dorsal
body wall to produce caucareous structures and the exclusive feeding structure,
the radula (BRUSCA & BRUSCA, 2002).
Gastropods are characterized by the
dorsal placement of viscera, cephalization and posterior location of the mantle
cavity (BRUSCA & BRUSCA, 2002). However, the systematic of gastropods is
not well developed and the groups still defined by the traditional orders
arrangement (RUPPERT
et al., 2004).
L.
filosa is part of the Litorinoidea super
family (intertidal periwinkles) which is part of the sub order Mesogastropoda defined by gastropods
with monopectinate gills, one atrium, one nephridium, simple osphradium and
presence of penis. Part of the order Caenogastropoda
wich includes the mesogastropods and the neogastropos (sub order Neogastropoda). The principal feature of
this order is the monopectinate gill that might help the expansion of this
animals to terrestrial ambient (RUPPERT
et al., 2004).
Due to distinctive characteristics of the tree
living periwinkles (Littoraria), they
were classified as Littorinopsis that
was lastly raised to a generic rank by Kuroba & Habe (1952) as cited by
Reid, 1986. Several discussions and reclassifications were proposed basing the
classification in the characteristics of the palial oviduct, sperm nurse cells
and penial shape until the Reid’s work been published in 1986 proposing a
summarized phylogeny. The lack of fossil records and information about ontogeny
makes the discussion about the genera more complicated and based in
geographical distribution (REID, 1986).
According to Reid et al. 2009, the estimated age of genus Littoraria is around 49-64 Ma, cf. New
molecular analysis showed better resolution, and the data for Littoraria have supported the previous
classifications ( REID et al., 2012).
The subgeneric studies about the Littoraria genus have confirmed the
genus as a monophyletic group supported by recognized synapomorphies. The
ovoviviparous species are allocated in the subgenus Littorinopsis where the L.filosa
was placed (REID, 1986).
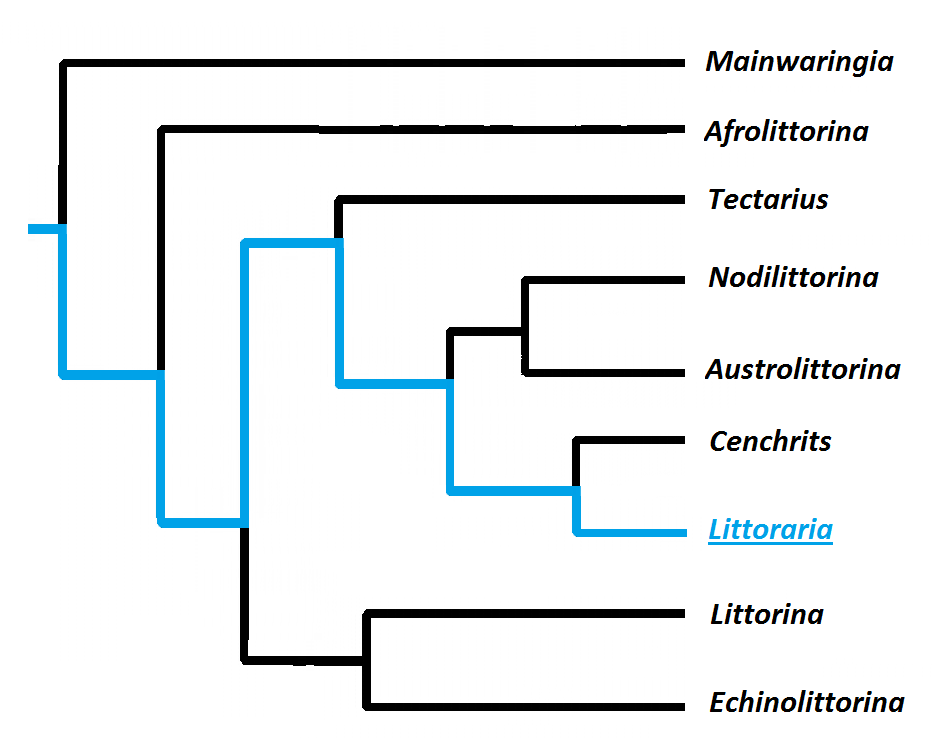 Figure: Cladogram representing the phylogeny of Litorinids. Littoraria (Blue). Gustavo Zoppello Toffoli, 2015 with reference to
Reid et al. 2012.
Figure: Cladogram representing the phylogeny of Litorinids. Littoraria (Blue). Gustavo Zoppello Toffoli, 2015 with reference to
Reid et al. 2012.
|
|
|
Biogeographic Distribution | |
The
Littoraia group is distributed only
in the tropical areas with just some representative species in subtropical areas.
Interestingly this confined position is not related with distribution of
mangroves since it is broad distributed in subtropical regions (REID, 1986).
Apparently, there are two endemic
centers, one in Malayan Peninsula, eastern Sumatra, western Borneo and southern
Vietnam where the richness achieve the top and a second one in the Australian’s
northeast coast where L.filosa is
found (REID, 1986).
The pattern of distribution found in
the Australian species may suggest competition. The L. luteola is largely found in New South Wales but rare in
Queensland where L. filosa is
abundant but rare in New South Wales. Furthermore, is known that in pacific
regions that leaf dwellings specie occurs, it occurs alone. This gradual
distribution showed in the eastern coast of Australia is a pattern found in
continental situations (REID, 1986).
In Littoraria the dispersion showed not related with the development
characteristic comparing oviparous and ovoviviparous species, instead of the
developmental features, the location apparently is more related with specie
dispersion since the planktonic eggs or larvae must travel in the ocean
currents to achieve new locations. Then, is noticeable that oceanic species
have more access to currents than continental species justifying their widely dispersion
(REID, 1986).
Individuals from southeast coast
shows differences in their shells when compared with the northeast ones, this
divergence is called clinal variation and is presented by F. filosa and F. articulate
individuals (REID, 1986).
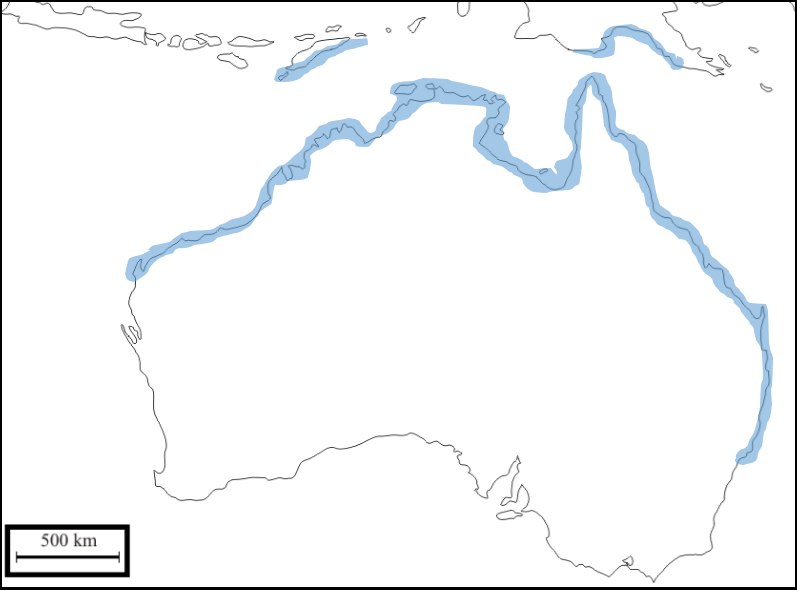 Map: Distribution of Littoraria (Littorinopsis)
filosa. Blue areas represents the
known occurrence of the species according to Reid et.al, 2009. Gustavo Zoppello Toffoli, 2015.
Map: Distribution of Littoraria (Littorinopsis)
filosa. Blue areas represents the
known occurrence of the species according to Reid et.al, 2009. Gustavo Zoppello Toffoli, 2015.
|
|
|
Conservation and Threats | |
Humans
do not use L. filosa as a food source
or for any economic activity, then this specie is more affected by the impact
that humans do in their habitat, the mangroves.
Mangroves
are water level sensitive environments that might change its pattern of
distribution according global changes that can affect the sea level,
temperature and climate patterns (REID & WILLIAMS, 2009). Due to the exclusive association with mangrove
trees (A. marina), L. filosa abundance and distribution
will be affected by any kind of disturb that might affect the mangrove
environment.
Mangroves are known as tropical and subtropical
halophytic plants that occurs along the coast working as nursery habitats and
as a shoreline shelter (DOWNTON, 1982). The grey mangrove A. marina is the dominant specie in Moreton Bay (MANSON et al., 2003), which is greatly impacted
by urbanization, causing stress to the mangrove habitat (POH, 2013).
Therefore, due to the dependence of L. filosa to the mangrove habitat,
actions in order to promote the conservation of mangroves will also promote the
conservation of the periwinkle. Is important to remember that conserving the
mangrove habitats, not just the periwinkles will be beneficiated but all the
other species that depends on the mangrove environment will be beneficieted
what can even influence the maintenance of the shorelines and fishery.
 Photo: Mangrove habitat at Esplanade Reserve. Gustavo Zoppello Toffoli, Wellington Point - Queensland
Photo: Mangrove habitat at Esplanade Reserve. Gustavo Zoppello Toffoli, Wellington Point - Queensland, 2015
|
|
|
References | |
BRUSCA, R. C., BRUSCA, G. J., 2002.
Invertebrates. 2nd Edition – Sinauer Associates
HICKMAN, JR, C. P., ROBERTS, L. S.,
KEEN, S. L., LARSON, A., L’ANSON, H., EISENHOUR, D. J., 2004. Integrated
principles of zoology. 12th Edition McGraw-Hill Higher Education
JOHANNESSON, K., EKENDAHL, A.,
2002. Selective predation favouring cryptic individuals of marine snails (Littorina). Biological Journal of the Linnean
Society (2002) 76: 137-144
MCKILLUP, S. C., MCKILLUP, R. V.,
2002. Flies that attack polymorphic snails on coloured backgrounds: selection
for crypsis by a sarcophagid parasitoid of Littoraria
filosa. Biological Journal of the Linnean Society (2002) 77: 367-377
MCKILLUP, S. C., MCKILLUP, R. V.,
2007. Apositary and selection for crypsis in the marine snail Littoraria filosa: an explanation for a
balanced colour polymorphism. Biological Journal of the Linnean Society (2008)
95: 62-71
PARSONAGE, S., HUGHES, J., 2001.
Natural selection and the distribution of shell colour morphs in three species
of Littoraria
(Gastropoda:Littorinidae) in Moreton Bay, Queensland. Biological Journal of the
Linnean Society (2002) 75: 219-232
REID, D. G., 1985. Habitat and
zonation patterns of Littoraria
species (Gastropoda: Littorinidae) in Indo-Pacific mangrove forests. Biological
Journal of the Linnean Society (1985) 26: 39-68
REID, D. G., 1986. The littorinid
molluscs of mangrove forests in the Indo-Pacific region. British Museum
(Natural History), London
REID, D. G., 1987. Natural
selection for apostasy and crypsis acting on shell colour polymorphism of a
mangrove snail, Littoraria filose (Sowerby)
(Gastropoda: Littorinidae). Biological Journal of the Linnean Society (1987)
30: 1-24
REID, D. G., DYAL, P., WILLIAMS, S.
T., 2012. A global molecular phylogeny of 147 periwinkle species (Gastropoda,
Littorininae). Zoologica Scripta 2012 – The Norwegian Academy of Science and
Letters, 41, 2, March 2012, pp 125-136
RUPPERT, E. E., FOX, R. S., BARNES,
R. D., 2004. Invertebrate zoology: a functional evolutionary approach. 7th
Edition – Thomson Brooks/Cole
|
|
|
|
|