Life History & Behaviour
Feeding
T. gratilla like most Echinoidea feed using the unique and highly developed protrusible jaw apparatus known as Aristotle's Lantern. This feeding apparatus is comprised of a set of 5 large calcareous plates which form pyrimid like structures joined together by transverse muscles. On the internal side of these plates are calcarious tooth structures, one for each plate (Ruppert et al. 2004). These teeth come together at the oral end of the lantern to form one beak. Specialized muscles for both protruding and retracting the teeth as well as opening and closing mechanisms, allow the urchins to pull, tare and scrape their desired food into the oral opening. Once the food has been taken into the lantern, it enters the buccal cavity, and pharynx which is joined to the esophagus. From here the food enters the stomach and begins its journey through the digestive tract and eventually being expelled out the anus on the oboral side of the urchin (Ruppert et al 2014).

Figure 1:Aristotle's lantern exterior diagram including the distal tooth for feeding.
(Ruppert et al. 2004)
T. gratilla have a diet that is comprised mainly of algal and other marine plants such as seagrass. Constantly grazing T. gratilla are continuous grazers feeding all throughout the day night cycle (Lawrence & Agatsuma 2007). Using their specialized Aristotle's Lantern they scrape, pull and tare at the algal bloom and seagrass, showing preference for different plant species depending again the geographical location (WoRMS 2014).
Locomotion
Tube feet are tentacle like structure which protrude out the test of the urchin alongside the spines. They extend their tube feet to grip onto substrate locking in place with the adhesive disc at the end of the feet and release via excrete a substance which releases the foot (Ruppert et al 2004). By using this process they are able to move across the substrate using any ambulacural area to lead, with no turning necessary to change direction of movement. The movement of these tube feet is driven by the water vascular system, which the water is pumped to the feet to enable extension and retraction. The water is pumped to a bulb which squeezes and pushes fluid into the tube feet (Ruppert et al 2004). The spines of the urchin also assist the urchin movement, although are not the primary movement organ, assisting in raising and falling of the body as well as aiding and horizontal movement (Lawrence & Agatsuma 2007).
 Figure 2: Tube feet extending for movement also the base of a
Figure 2: Tube feet extending for movement also the base of a
container during an observation of T. gratilla movement
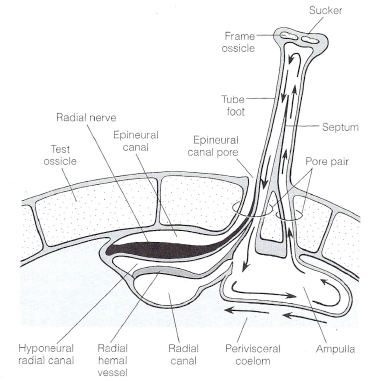 Figure 3: Respiration and direction of water movement inside the tube feet (Ruppert et al. 2004)
Figure 3: Respiration and direction of water movement inside the tube feet (Ruppert et al. 2004)
Tube feet are not only the means of locomotion but are also gills the supply water to the water vascular system and the perivisceral cavity. Figure 3 outlines this tube feet respiration, the fluid streams are separated by a septum located in the urchins foot which prevents the diffusion between the O2 rich stream (left side of the foot) and the O2 depleted stream on the right (Ruppert et al 2004). Like most urchins T. gratilla have 5 pairs of peristomial gills on their peristomial membrane, each being a highly branched out-pocket of the peripharyngeal coelom. Muscles and ossicles associated with Aristotle's lantern pumps coelomic fluid into these gills. It is believed that these peristomial gills are the surface for gas exchange for the lanterns musculature (Ruppert et al 2004).
Reproduction
It is difficult purely on observation to distinguish between sexes of T. gratilla, as they do no display any external indication of sex. As gonotrophic (remain the one genender i.e one of two sexes) organisms T. gratilla must go through sexual reproduction in order to reproduce. Reproducing occurs via spawning when conditions are optimum which are given off by certain environmental indicators (Lawrence & Agatsuma 2007). When conditions are correct a mass spawning even occurs, where gametes from both sexes are expelled into the water column (Lawrence & Agatsoma 2004). Sea urchins have been observed to gather together in close proximity before these events occur, it is believed that they gather together to increase the chances of fertilization before gametes begin to deplete. Once fertilized cleveage begins in and at the eight cells stage the blastula ciliated and begins to form 4 small micromeres (Ruppert et al 2004). During larval development, four paris of arms supported by internal skeletal rods. The larvae can swim up to months feeding in the water column before finally settling on the ocean floor (Ruppert et al 2004).
In order to gather more information on the reproductive cycle of the urchin a study was conducted to determine the effects of ocean acidification from flash flooding on T. gratilla development.
Tripneustes gratilla are commonly found on ocean floors of coastal regions preferring to settleon rocky substrate such as coral and man-made reefs. Due to their close proximity to land they are often subject to anthropogenic disturbances. Rarely aware of the affects man can have on such unfamiliar species of marine invertebrates, as society is often only drawn to damages to fellow humans and other well known larger vertebrates (Donpont et al 2010). Everyday chemicals and inorganic substances are washed down the waterways from cities and towns and enter marine ecosystems like the Moreton Bay area (Caldeira & Wickett 2010). Moreton Bay is home to a population of T. gratilla, and similar numbers are found along the Great Barrier Reef off the east coast of Queensland, Australia (Lawrence & Agatsuma 2007). The Great Barrier reef has around 23 major rivers flowing into the same region. Flooding events lead to changes in the composition of the oceanic waters, where chemicals such as fertilisers are washed down stream and can quite easily cause rapid acidification of the typically basic ocean waters. Most marine species are sensitive to this and T. gratilla is no different (Caldeira & Wickett 2010). As spawners they rely on the ocean conditions to not only trigger spawning but allow for the survival of the gametes and the fertilised zygote (Lawrence & Agatsuma 2007). Therefor slight changes in these conditions could theoretically alter the survival and fertilisation rates of this species. This prediction is the aim of this experiment, hypothesizing that the more acidic the water becomes, the rate of fertilisation within the species will deplete.
Methods
Using a generic fertiliser that is found in rural and urban areas 3 different levels of pH were determined. The control had no fertiliser added and only fresh seawater from Manly in Moreton Bay the pH was checked to ensure it was on par with the generalized ocean pH level (≤ 8.5). The other two were measured to 8.0-7.0 and 7.0-6.0 respectively, giving the modified slight and extreme disturbances in the water.
This study was conducted by using 2 specimens (1 male and 1female), specimens were collected from a passage between Amity point on North Stradbroke Island and the south end of Moreton Island. They were transported back the University of Queensland and held in an artificial tank for an extended period of time with conditions to match natural circumstances. Using a number of larger individuals, the specimens were selected at random and injected with around 2.0mL of 0.5 M KCL into the column. The specimens were then shaken and placed back into the water leaving the aboral opening out of the water. If a white liquid began to excrete out the gonophore, the individual is male and the sperm was pipetted into a tube, careful not to get it wet. If the substance was yellow to individual was placed on top of a beaker full of seawater with the gonophore facing down so that the eggs fall to the bottom.
 Figure 4: Experimental set up with the eggs in the beaker on the left and the three pH level beakers going from 8.5
Figure 4: Experimental set up with the eggs in the beaker on the left and the three pH level beakers going from 8.5
(control) on the left and the altered two on the right.
Approximately 1mL of egg mass were pipetted into each of thethree beakers, the sperm was the diluted down to 10 microliters to 100mL, 1mL from this sperm dilution was added to each of the beakers at 5 minuet intervals to begin fertilisation The beakers were counted for fertilisation every 10 minuets using a cavity slide where the number of fertilised eggs were counted from left to right until the whole slide is covered. This was repeated two more times to ensureaccurate estimates of the number of fertilisations. After the final count the water in the beaker was discarded and replaced with the same level of pH ineach. Using the same sperm and egg the fertilisation experiments were carried out two more times. A statistical analysis on the results was then conducted to determine the significance of theresults.
Results
The results showed that fertilization rate in the control experiment is much higher than the 8.0-7.0 and 7.0-6.0 experiments. In all 3 of the trials the control had high amounts of fertilization while 8.0-7.0 had very few and 7.0-6.0 had no fertilizations occurring at all. An ANOVA test was conducted to determine the statistical significance which showed p<0.05 making the results statistically significant based on the ANOVA.
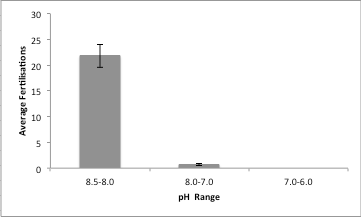 Figure 5: Chart displaying the average fertilisation rate for the three sets on pH levels over the
Figure 5: Chart displaying the average fertilisation rate for the three sets on pH levels over the
three replicates including standard error bars. Using an ANOVA a p value of p<0.05 was obtained
Discussion
It was expected that changing the pH of the water would alter the number of fertilizations in each of the replicates, this was reflected in the data. When the pH ranged from 8.5-8.0 the fertilization rate was quite high with an average around 22 per count, 8.0-7.0 has a substantially lower rate with an average of .66 and the final range 7.0-6.0 having no fertilizations counted in all three of the replicates. As previously stated this pattern was expected, however not the the extent of the results gathered. The results similar study conducted on the same species focusing more on the larval development when temperature and pH was altered. The study conducted experiments on the effects of altering these environmental conditions while the species is in its planktonic larval form. Sheppard & Brennand et al. (2010) found the when the pH and temperature increased the larvae began to show signs of dwarfing effects, which can negatively impact on adults benthic communities by lowering the fitness of the organism and its ability to compete for resources (Sheppard Brennand et al. 2010). The levels in which the pH was altered in Sheppard & Brennand et al. (2010) were on a much narrower scale than the ones used in this study due to lack of availability to high quality test kits. However if this study was done on a much broader scale with greater exposure to resources a greater indication of the effects of flooding and ocean acidification can be obtained. Exposure of this information is vital to not only ensure that these disturbances have smaller impacts on this species but other marine invertebrates which serve a greater cause to the function of the ecosystem then most of the population is aware of.
|