Life History & Behaviour
Feeding and Digestion
In situ observations have shown the use of a spherical mucous web, which is deployed to capture prey items (Newman 1990). In this way, thecosome pteropods are analogous to terrestrial spiders, using a 'trapping' mechanism for feeding. Studies on other species within the family Cavoliniidae, including the closely related C. virgula, have shown that when the web is present the individual hangs without flapping the wings, which instead are folded dorsally. This suggests the bilobed mucous web may also assist with buoyancy (Gilmer & Harbison 1986).
It was traditionally thought that C. acicula fed using a ciliary feeding mechanism, in which food is passed from the ciliated fields (cf) situated on the ‘wings', to the mouth (m). This ciliated current was described by Yonge in 1926, which was observed by placing specimens in shallow glass dishes containing seawater and adding drops of coloured carmine to observe currents. Gilmer and Harbison (1986) noted that the ciliary beating is not involved in food capture, but rather the deployment and ingestion of the mucous web. As the body and shell are transparent, it is possible to follow the course of food through the mouth and the length of the gut (click here to watch a video).
From the mouth, food is passed into the globular buccal mass (bm) which contains a small radula with around ten transverse rows of three teeth. Food is passed into the oesophagus (oes), from which a swallowing motion can be distinctly observed. The ‘stomach’ is described by Pelseneer (1888) as a“masticatory gizzard with muscular walls… four large symmetrical plates, a fifth posterior plate and generally eight small anterior plates”. Behind the gizzard is a chamber with the digestive gland (liver), which usually yellow or brown and is modified to accommodate the needle-like shape of the body. The digestive gland of freshly caught animals is often greenish brown, which reflects their generalist diet of diatoms, dinoflagellates, radiolarians and foramniferans. Prey size is restricted to the maximum dimensions of the ciliated tracts on the wings (Newman 1990). The anus (a) empties on the left into the pallial cavity. Digestive wastes are compacted into strings and channeled out over the dorsal surface of the mantle (Newman 1990). These strings are fragile and easily detached, and can attract swarms of copepods (Newman 1990; Gilmer & Harbison 1986).
Reproduction
All pteropods in the family cavoliniidae are protandric hermaphrodites, which means that they develop from males into hermaphrodites and then into females (Newman 1990). Sexual maturity is usually reached withina year. Veligers hatch from egg masses which remain planktonic. Differentiation of juvenile cavoliniids is difficult, as the shells look similar and can differ from their adult forms (Newman 1990).
C. acicula veligers have a ciliated velum (Figure 1), a four-lobed structure that is used for both swimming and food collection. Click here to watch a video that shows a cavoliniidae veliger, from the genus creseis, swimming using the ciliated lobes rather than the characteristic ‘flapping’ of adults.
Bandel and Hemleben (1995) described the reproduction of C. acicula in detail, with laboratory observations on mature individuals caught in the Gulf of Aden. Eggs were 0.06-0.07mm and were surrounded by a thin mucous sheath. Each individual produced 110-130 fertilised eggs. Development of the embryo initially resembles the characteristic trochophore larvae within the egg capsule, which hatches into a veliger after approximately 60 hours of development. Newborn veligers can completely retract into their tubular shells,which are around 0.1mm long and resemble the adult form.
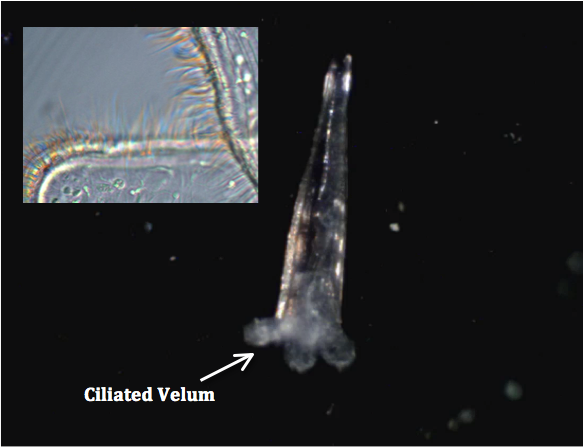
Figure 1: Creseis veliger, with ciliated lobe detail.
Locomotion
All pteropods have wing-like appendages (pteropod = ‘wingfoot’) which they ‘flap’ for forward locomotion (Figure 2). Gilmer and Harbison (1986) reported that C. acicula does not swim or move at all whilst feeding and swimming is only used an escape response.
In observations conducted on specimens collected from Heron Island in the Great Barrier Reef, individuals were observed swimming from the bottom of 20cm deep glass jars in an upward motion, by rapidly flapping their wings. This movement was also recorded on specimens in shallow petrie dishes, and can be seen in this video (click here). The shell was maintained in a vertical position, and ‘flapping’ occurred in short intervals, followed by periods of rest. Whenever motion ceased, C. acicula would slowly descend with wings stretched out, a behaviour also recorded by Kornicker (1959).
Most thecosome pteropods exhibit diel vertical migrations, in which they move to deeper layers of the water column during the day and rise to the surface at night. A study by Gilmer and Harbison (1986) on the closely related Creseis virgula showed evidence of negative phototaxis, as individuals responded to a light source by drawing in the feeding web and sinking at approximately 10cms-L. Rate of descent is likely controlled by the wings of the animal, making adjustments to angle, pitch and horizontal breadth of the outstretched lobes (Kornicker 1959).
Figure 2: Adult C. acicula with extended wings.
Respiration
Respiration is thought to occur over the wing surface in the majority of species, though no specific studies have been conducted on C. acicula (Newman 1990). An experiment by Gilmer (1974) on pteropod respiration showed a common metazoan trend, with oxygen consumption decreasing per unit weight, with an increase in size.
|