Life History & Behaviour
Feeding
C. tramoserica is an opportunistic forager; feeding only when conditions are suitable and preferably being submerged under the tide (Gray& Williams, 2010). They hardly refuge on the rocks with macroalgal growth due to difficulty of adhering on the substrates. The animal gazes to the film of microalgae on the rock surface; remains algal spores and microalgae in cracks and crevices where they are unlikely to reach (Underwood & Jernakoff, 1981).
Reproduction
C. tramoserica is a sexual reproduction with eggs and sperm being broadcast into the sea. Spawning usually occurs in summer season but also correlated with the site. There are less spawned populations from January to March at the middle intertidal but more in low and high intertidal zone (Fletcher 1984). From June to September, larval stages developed over this time when food availability becomes abundant (Fletcher, 1984). Juvenile growth is variable between age groups, but on average the size increased from 6 to 32 mm in about two and a half years. Sexual maturity is reached at an age of one year, when shells are 24 mm in length (Parry, 1982).
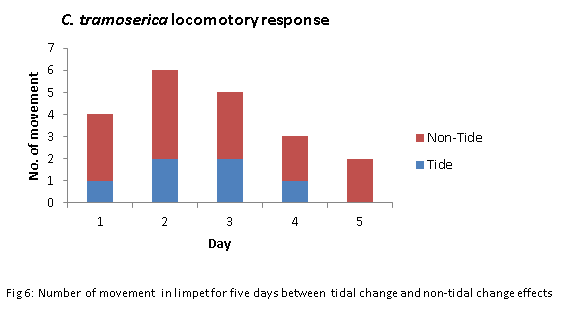
Locomotion Experiment
Some limpet species have unique locomotory activity as they recognise their ‘home’ and return to the same spot after the feeding cycle from distance by a recognition of mucus tracks secreted on the rock (Underwood, 1977). Some species move randomly after days of feeding and end up away from the starting point as a new home (Mackay & Underwood, 1977). The various homing behaviours are believably regulated by local density and dispersion to maximise utilisation of the algal food resource.
A design experiment is conducted to further establish whether the rhythms of locomotor exhibited by C. tramoserica on the shore is caused by particular foraging patterns, tide changes or other stimulations from the environmental fluxes such as the thermal effect.
Method
Two groups of 5 specimens were randomly collected for experimental and control test during the low tide (0.8m) of the shore. They were carefully removed by a knife and brought back for experimental setups. The first group was placed in a 30 x 19 x 17(cm) aquarium tank and fed by microalgae attached on the rocks originate collected from the field. The tank was placed in semi-outdoor area so that the photosynthetic process of algae could proceed. The water was also replaced twice a day to maintain the quality of biomass and bacteria available to the limpets and other animals. The limpets also underwent the tidal change by regularly removed and added seawater in the tank twice a day and sprayed the water closely on the shells as pretending to be awashed by the wave action (Gray & Williams, 2010). The control specimens were placed in the same experimental setups without the tidal effect so that they will be submerged in the water throughout the experimental process. The movement pattern of limpets was measured by counting the number of limpets that had any distance changes within 1 hour of the tidal rised and fell in five days. All the specimens, animals and rocks were returned to the original area after completion of the experiment.
The null hypothesis predicts the mean of two groups in their locomotory patterns is the same. The alternative hypothesis considers a significant difference in locomotory responese between the tidal and non-tidal effects. A significant value of P is <0.05. The statistical method was as non-parametric Mann-Whitney one-tailed t test.
Result
The result of hypothesis test in fig.1 indicated as 0.024 which was less than P value <0.05. It concluded that a significant difference in the moving pattern between tidal and non-tidal effect.
Fig 4: Mann-Whitney hypothesis result in SPSS. Significant t-test result = 0.024
|
Test Statistics
|
|
Limpet
|
|
Mann-Whitney U
|
2.000
|
|
Z
|
-2.278
|
|
Exact Sig. (2-tailed)
|
.048
|
|
Exact Sig. (1-tailed)
|
.024
|
|
Point Probability
|
.024
|
In addition, the mean rank in fig 2 shown that the limpets with tidal impacts was twice lower than the limpets with non-tidal impacts (3.4 <7.6). It indicated a less movement being occurred in the cause of the tidal change compared with the non-tidal change environment. The most active limpets in both two groups were in day 2, followed by day 3. The only movement occurred in day 5 was the non-tidal change group (fig. 6).
Fig 5: the mean rank comparison between Group 1 (3.40) and Group 2 (7.6)
|
Ranks
|
|
Group
|
N
|
Mean Rank
|
Sum of Ranks
|
|
Limpet
|
1
|
5
|
3.40
|
17.00
|
|
2
|
5
|
7.60
|
38.00
|
|
Total
|
10
|
|
|
Group1=Tidal effects Group 2=Non Tidal effects
Discussion
Foraging Behaviour
Factors causing of locomotory response in C. tramoserica are various. The rhythmic foraging behaviour involved maximising the grazing opportunity when feeding competition occurred due to density increased in the populations (Branch & Branch 1980). An observation from the second group found that two closed refuge limpets had moved apart after two days of the experiment. The film of algae on the rocks was also reduced throughout the 5 days experiment which assumed some grazing activities taking place regardless the tidal changes to affect their locomotory responses.
Wave Action
The factor of wave actions causing aggregated movements by the tidal simulation is also predicted. From the first group, 3 out of 5 limpets moved up to the tank wall and remained intact for three days where no tidal was reached. A reason of this is because unfavourable environmental conditions can be avoided by achieving in group actions than individuals (Coleman et al, 2004). It is suggested that this aggregation moving pattern prone to water retention on the surface of rock and airflow between the limpets compared with solitary individuals Coleman (2010). The rising seawater allows the limpets to take water in the mantle to prevent from desiccation.
Thermal Stress
Thermal stress varies in the intertidal zone over multiple spatial and latitudinal scales (Morley et al, 2012) considerably affect the locomotion pattern between emersion and immersion through each tidal cycle. A temperate limpet C. tramoserica have lowest thermal safety margin (TSM) compared with the tropical species such as C.radiate, which presumably would be less physical response required during the tidal change. This considerbly indicated from the group 1 result where the limpets required less movement and water submerge required. However, the thermal factor is not a scope of this experiment as no related data to be collected. Therefore, the further statistical analysis in proving the contribution of locomotion based on thermal stress is needed.
The grazing movement of C tramoserica (video): http://youtu.be/dfJ-IX6njMk
|